 2013, Vol. 34
2013, Vol. 34文章信息
- 张安杰, 曹振东, 付世建
- ZHANG Anjie, CAO Zhendong, FU Shijian
- 生境不完全重叠的两种鲤科鱼类耐低氧及运动能力比较
- Comparison of hypoxia tolerance and locomotor performance in two cyprinids with incompletely overlapped habitat
- 生态学报, 2013, 34(20): 5860-5867
- Acta Ecologica Sinica, 2013, 34(20): 5860-5867
- http://dx.doi.org/10.5846/stxb201301230130
-
文章历史
- 收稿日期:2013-1-23
- 网络收稿日期:2014-3-11






不同河流生境存在着巨大的环境差异,主要表现在温度、溶氧水平和水流速度等方面。河面的宽度和深度对溶氧水平和水流速度均有影响。鱼类的低氧耐受能力、身体形态和游泳能力等均受生境(温度、水流和溶氧等)变化的影响[1, 2]。尽管有研究者提出富营养化和温室效应是环境低氧的主要原因[3],但在河流中水流速度同样也是一个形成低氧环境的因素;相对静止水体的溶氧水平低于水流湍急的河流或小溪流的溶氧水平。耐低氧能力越强的鱼类在低氧环境中能更好的生存、繁殖和生长[4, 5, 6, 7, 8]。已有研究分别以临界氧分压 (Pcrit)[4, 5]、水面呼吸 (ASR)[6]和失去平衡点 (LOE)[7]等指标评价鱼类的低氧耐受能力。Pcrit、ASR和LOE越低的鱼类低氧耐受能力越强[4, 5, 7],其中Pcrit和LOE侧重于生理反应,而ASR则主要侧重于行为反应[6]。已有研究报道,在溶氧水平较低的环境中生存的鱼类具有较强的低氧耐受能力[1, 8]。 水体溶氧水平决定着鱼类的生存,同时对鱼类的运动能力和行为特征产生影响[9, 10, 11]。鱼类的游泳运动能力与其逃避敌害和成功捕食密切相关[12];通常用临界游泳速度 (Ucrit) 作为鱼类有氧运动能力的主要评价指标[13]。且与其生存环境的水流速度有关,在湍流环境中生存的鱼类与缓流或水流相对静止的环境中生存的鱼类相比具有较强的游泳运动能力[2]。同时,鱼类的Ucrit也受到环境低氧的影响,且随着溶氧水平的下降而降低[14, 15, 16]。运动耗氧率 (MO2) 作为评价鱼类游泳过程中能量消耗的重要指标,同样也受到环境溶氧水平的影响[14, 15]。因此,环境水流速度较高对鱼类的运动能力要求较高,但该环境溶氧水平较高,对其低氧耐受能力要求则较低;在水流速度较低或相对静止的环境中生存的鱼类需要有更强的低氧耐受能力,但对其运动能力的要求则相对较低。 乌江作为长江上游右岸最大的支流具有丰富的环境多样性,即使是在同一河段的不同地点也存在水流速度、溶氧水平等生境的差异。马口鱼 (Opsariichthys bidens) 和宽鳍鱲 (Zacco platypus) 虽然是乌江流域分布广泛且亲缘关系近的两种小型鲤科鱼类,但通过最近几年对乌江流域的考察发现,在水流速度快的上游或下游小型支流中均有宽鳍鱲分布,但在这些河流中未见马口鱼的分布。即使在马口鱼和宽鳍均有分布的河段,但其仍有各自所偏好的生存环境,马口鱼更偏好水流缓慢或相对静止的水体中生存,而宽鳍鱲则更偏好水流湍急的水体中栖息。因此,这两种生境不完全重叠的鱼类也许会出现不同的低氧耐受能力和在不同溶氧水平下可能会表现出不同的生理反应。据此本研究选取马口鱼和宽鳍鱲作为研究对象,在(25.0±1.0) ℃条件下,分别测定进行低氧耐受能力 (Pcrit、ASR、LOE) 和低氧 (8、4和2 mg/L) 运动能力 (Ucrit、MO2) 的测定。考察并比较两种实验鱼的耐低氧能力和低氧下的运动能力及其代谢特征,以及两种实验鱼生理功能及其机制的差异与生存环境和生活习性的关联。
1 材料方法 1.1 实验鱼来源与驯养于2012年5月初,位于乌江下游羊角镇 (N29°23′,E107°36′) 江段右岸岸边水流速度缓慢且有回水的地方捕获马口鱼 (N=40,体长5.3—12.5 cm),在右岸一条坡降大且水流湍急的小溪流捕获宽鳍鱲 (N=40,体长 5.1—7.8cm)(图 1),置于自净化驯化控温循环水养殖系统驯养两周。驯养水温为(25.0±1.0) ℃,驯养水体溶氧水平 > 7.0 mg/L,每日用充分曝气后的自来水置换水体总水量的10%,光周期为12 L ∶ 12 D。驯养期间每天9:00以商业颗粒饵料饱足投喂1次,投喂半小时后清除残饵和粪便,每组实验鱼在实验前禁食48 h。

|
| 图 1 乌江下游马口鱼 (Opsariichthys bidens) 和宽鳍鱲 (Zacco platypus) 的捕获地点 Fig. 1 The collecting site of hook snout carp (Opsariichthys bidens) and pale chub (Zacco platypus) at the lower reaches of Wu River |
| 种类pecies | 水体温度Temperature/℃ | 水流速度 peed/(m/s) | 溶氧水平Dissolved oxygen/(mg/L) | 河道宽度 Wide/m |
| 马口鱼Opsariichthys bidens | 18.9 | <1 | 8.88 | > 100 |
| 宽鳍鱲Zacco platypus | 20 | > 3 | 10.3 | <5 |
在温度驯化两周后选取体重、体长相近的实验鱼,在(25.0±1.0) ℃条件下分别进行低氧生存耐受能力和运动耐受能力的测定。以Pcrit、ASR和 LOE 3个不同指标考查低氧生存耐受能力;分别在不同溶氧水 (8、4和2 mg/L) 下测定实验鱼的Ucrit及其运动MO2,将运动耗氧率的最大值称为活跃耗氧率 (MO2active)。
1.3 测定方法及主要参数 1.3.1 ASR和 LOE的测定采取氮气置换法将水体溶氧浓度阶段下降的方式改变实验水环境 (120 L圆柱形水体)。溶氧梯度分别设置为5、2.5、1.2、1.1、1.0、0.9、0.8、0.7、0.6 mg/L(±0.02 mg/L) 每个溶氧水平调整时间和维持稳定时间均为0.5 h左右。在实验开始前将实验鱼分别转移至实验水体适应24 h,在适应期间实验水体溶氧接近饱和。在实验过程中每间隔3 min观察并记录出现ASR或LOE的实验鱼数量,该实验进行3个重复,每个重复有实验鱼 (n=4)。根据每个重复所记录的实验鱼数量计算50% ASR和50% LOE,具体计算方法如下:
式中,[O2]为各观察点的溶氧值,[O2]0是实验鱼出现水面呼吸或失去平衡的前一个溶氧梯度,t为实验鱼在某个溶氧浓度下出现水面呼吸或失去平衡的历时,Δ[O2]为实验鱼在失去平衡时的溶氧浓度和前一个溶氧浓度的差值。 式中,ASR表示水面呼吸百分比,R表示在每一个观察点出现ASR的实验鱼条数。R0为该观察点的实验鱼总条数。然后根据实验鱼每个重复的ASR百分比和LOE百分比与该观察点溶氧值 (通过公式1得到每个观察点的溶氧值) 进行拟合,最终计算得到每个重复的50% ASR和50% LOE溶氧值。 1.3.2 不同溶氧水平下的静止耗氧率 (MO2rest) 与Pcrit的测定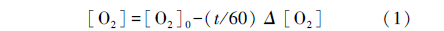
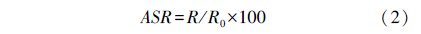
采用密闭式呼吸代谢仪测定每条实验鱼的MO2rest和Pcrit,将每条实验鱼均转移避光的有机玻璃管中适应24 h,在适应期间,各呼吸室开启并用循环水泵供水,以确保呼吸室内溶氧水平接近饱和。开始测定时,呼吸室关闭并使用溶氧探头 (HQ20,Hach Company,Loveland,Colorado,USA) 每间隔3 min连续检测溶氧水平的变化,直到实验鱼出现失去平衡的现象时结束实验。不同溶氧水平下的静止耗氧率和临界氧分压的计算方法如下:
式中,[O2]k (μmol/L) 为取样时间点k时的氧浓度;[O2]k+1 (μmol/L) 为下一个时间点的浓度 (根据在相应的温度和压强下水中的O2溶解系数来计算这些值);VOL (L) 为呼吸室的总体积减去鱼的体积;t (h) 为时间点k和k+1间的间隔;和m (g) 鱼的体重。MO2 (mg kg-1h-1)为时间点k+1的单位体重耗氧率[14]。将代谢率和溶氧水平作图,采用“双线法 (two-segmented straight lines)”求出MO2随溶氧下降而出现显著下降拐点,即为Pcrit[17]。 1.3.3 Ucrit和运动 MO2的测定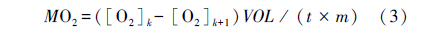
使用自行设计的密闭式鱼类游泳代谢测定仪进行实验测定,其结构与工作原理已在相关文献[18, 19] 中进行了详细介绍。实验开始前将游泳代谢仪浸没于长方形恒温水浴水体 (体积) 中,然后将实验鱼分别转入游泳管在水流速度为24.0 cm/s时适应 1 h,以消除转移和低氧环境胁迫的影响;随后采用国际通用方法[19]进行Ucrit 测定。测定采用24.0 cm/s为起始速度,以 6.0 cm/s为速度增量 (ΔU),以 20 min 为持续时间 (ΔT),游泳速度不断增加直至实验鱼达到运动力竭状态。力竭状态的评判标准为实验鱼停靠在游泳管末端筛板 20 s以上[19]。测定过程中每20 min换水1次,以确保密闭游泳管内的水体溶氧水平不因实验鱼的耗氧而低于初始水平的75%,使用溶氧仪每2 min连续监测1次溶氧水平的变化[20]。实验结束后将实验鱼取出,对游泳代谢仪重新密闭并进行20 min的耗氧率测定,以扣除细菌耗氧的影响[19]。为了满足在不同溶氧水平下Ucrit和运动 MO2的测定需求,将水浴水体上方覆盖塑料薄膜并用氮气置换水体溶氧的方法使其溶氧水平达到设定浓度 (8、4和2 mg/L)[20];测定方法如上所述。Ucrit和运动MO2的计算方法如下:
式中,Ucrit (cm/s) 表示临界游泳速度,U为实验鱼能够完成设定时间 (20 min) 的最大游泳速度,ΔU是速度增量 (6.0 cm/s),T为设定的持续游泳历时 (T =20 min),t为未能完成设定历时的实际持续游泳时间 (t<20 min)。 式中,MO2 (mg O2 kg-1h-1) 代表耗氧率,St为每档期间 (20 min) 溶氧值随时间变化的斜率,S0为空白组的溶氧值变化斜率,3.45为运动呼吸仪的系统体积 (L),W表示体重 (g)。 1.4 数据分析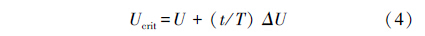
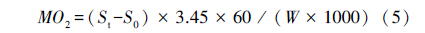
用Excel (2003) 和SPSS 17.0对所有实验数据进行常规统计和方差分析。所有数据表示为平均值±标准误 (mean±SE),差异显著水平为P<0.05。使用t-检验对两种实验鱼的Pcrit、LOE和 ASR进行差异显著性分析。利用双因素方差分析对种类和溶氧水平与Ucrit和 MO2active的关系进行分析。在双因素方差分析后通过Duncan鉴定平均值的差异显著性。
2 结果 2.1 马口鱼和宽鳍鱲的低氧耐受能力比较马口鱼的Pcrit (2.44±0.20) mg/L 显著高于宽鳍鱲 (1.86±0.10) mg/L(P=0.031)(图 2)。但马口鱼的LOE (0.84±0.01) mg/L 显著低于宽鳍鱲 (0.97±0.02) mg/L(P=0.004);马口鱼的50% ASR (1.23±0.16) mg/L同样显著低于宽鳍鱲 (1.97±0.11) mg/L(P=0.023)。

|
| 图 2 马口鱼和宽鳍鱲的临界氧分压 (Pcrit,平均值±标准误,n=7) 、水面呼吸 (ASR) 和失去平衡点 (LOE,平均值±标准误,n=12) Fig. 2 The critical oxygen tension (Pcrit) ,aquatic surface respiration (ASR) and loss of equilibrium (LOE) of hook snout carp and pale chub *表示差异显著 (P<0.05) |

|
| 图 3 溶氧对马口鱼和宽鳍鱲代谢率 (MO2) 的影响 Fig. 3 The effect of dissolved oxygen level on oxygen consumption rate (MO2) in hook snout carp and pale chub |
两种实验鱼的Ucrit均随着测定溶氧水平的降低而下降,但不存在交互作用 (P=0.208)。在各个溶氧水平下马口鱼的Ucrit均显著低于宽鳍鱲,并在测定溶氧浓度为8.0和4.0 mg/L时差异显著 (图 4)(P<0.01)。两种实验鱼的运动MO2均随着水流速度的增加而呈现指数增加,但随着测定溶氧水平的降低耗氧曲线变得相对平缓,尤其是在溶氧为4.0 mg/L时马口鱼的耗氧曲线与宽鳍鱲相比越发平缓 (图 5),表明马口鱼在运动状态下更早的启动无氧代谢。 两种实验鱼的MO2active 随溶氧水平的降低而下降,且存在交互作用 (P=0.005);并在测定溶氧水平为4.0和2.0 mg/L时差异显著 (图 6)(P<0.001)。

|
| 图 4 溶氧水平对马口鱼和宽鳍鱲临界游泳速度 (Ucrit) 的影响 (平均值±标准误,n=8) Fig. 4 The effect of different dissolved oxygen level on critical swimming speed (Ucrit) of hook snout carp and pale chub a,b,c,A,B,C不同上标字母表示马口鱼或宽鳍鱲分别在不同溶氧水平下差异显著 (P<0.05); *表示同一测定溶氧水平下马口鱼和宽鳍鱲游泳能力间差异显著 |

|
| 图 5 在不同溶氧水平和不同游泳速度对运动耗氧率 (MO2) 的影响 (平均值±标准误,n=8) Fig. 5 The effect of different dissolved oxygen level and swimming speed on oxygen consumption rate (MO2) in hook snout carp and pale chub |

|
| 图 6 溶氧水平对马口鱼和宽鳍鱲的活跃耗氧率 (MO2active) 的影响 (平均值±标准误,n=8) Fig. 6 The effect of dissolved oxygen level on active oxygen consumption rate (MO2active) in hook snout carp and pale chub a,b,c,A,B不同上标字母表示马口鱼或宽鳍鱲分别在不同溶氧水平下差异显著 (P<0.05); * 表示同一测定溶氧水平下马口鱼和宽鳍鱲游泳能力间差异显著 |
鱼类能否在低氧环境中生存取决于其低氧生存耐受能力的强弱,通常用Pcrit、LOE和ASR为其评价指标[4, 5, 6, 7]。本研究发现马口鱼的LOE和ASR均显著低于宽鳍鱲,然而马口鱼的Pcrit却显著高于宽鳍鱲 (图 2)。已有研究报道,由于Pcrit是一个以鱼类耗氧率随溶氧水平下降而变化来评价鱼类低氧耐受能力的指标,所以在测定过程中实验鱼的活跃性会导致Pcrit值被过高或过低估计[21, 22, 23];而LOE和ASR则是从神经系统和行为方面来评价鱼类的低氧耐受能力[24]。在本研究中,马口鱼在Pcrit点前的耗氧率随着溶氧水平的降低而呈现上升趋势,而宽鳍鱲则趋于稳定 (图 3);马口鱼和宽鳍鱲相比由于马口鱼更为活跃而导致其Pcrit被偏高估计。因此,本研究采用LOE和ASR作为评价马口鱼和宽鳍鱲低氧耐受能力的指标更为可靠。据此,本研究结果表明马口鱼与宽鳍鱲相比具有更强的低氧耐受能力。 乌江流域具有丰富的环境多样性,对于鱼类来说拥有足够丰富的生境可以供其选择偏好的环境生存。野外生存的这两种实验鱼在乌江流域的分布并不完全重叠,即使在同一地点的不同河段分布也不尽相同。已有研究发现,生活在沼泽地区鱼类比生活在开阔水域的鱼类具有更强的低氧耐受能力[1]。本研究发现,更偏好生活在缓流或相对静止水体中的马口鱼与更偏好生活在湍流环境中的宽鳍鱲具有更强低氧耐受能力,这可能就与它们所偏好生活环境的选择有关。
3.2 两种实验鱼在不同溶氧水平下的运动能力及其运动代谢鱼类的运动能力与其逃逸、捕食和繁殖等行为密切相关[12, 25, 26];除水温[27]、饥饿[28]和摄食水平[29]等对鱼类的临界游泳速度产生影响外,生存环境的水流速度同样对其运动能力产生显著影响[2]。在不同水流环境中生存的鱼类具有不同的运动能力,在水流速度较高的环境中生存的鱼类具有较强的运动能力[30]。在本研究中,湍流环境中生存的宽鳍鱲比生存在缓流或水流相对静止环境中生存的马口鱼的运动能力更强,且在测定溶氧浓度为8.0和4.0 mg/L时差异达到显著水平。因此,宽鳍鱲的运动能力强于马口鱼可能与它们所偏好的生存环境不同而至。 鱼类的游泳运动主要利用有氧代谢提供能量[12]。有研究报道,鱼类的临界游泳速度会随着溶氧水平的降低而下降[14, 15, 31],在本研究中马口鱼和宽鳍鱲的Ucrit从溶氧水平为8.0 mg/L到4.0 mg/L时分别下降了25.6%和17.9%,并且两种实验鱼的临界游泳速度随溶氧水平下降的表现一致。鱼类在运动过程中的能量消耗通常用临界游泳过程中的耗氧率作为评价其有氧代谢能力的指标[12, 32],当实验鱼在即将达到Ucrit时存在一定比例的无氧代谢的参与[12, 33, 34]。已有研究发现,在低氧条件下鱼类随水流速度增加而增加的耗氧曲线均表现出扁平或降低的趋势[14, 15, 20]。在本研究中,马口鱼和宽鳍鱲的运动MO2均随游泳速度的增加而呈现指数增加,但随着测定溶氧水平的降低耗氧曲线变得相对平缓(图 5);在测定溶氧水平为4.0 mg/L时,在高流速下马口鱼的耗氧曲线与宽鳍鱲相比扁平程度更大(图 5),说明马口鱼在相对更低的速度下就启动无氧代谢为其游泳提供能量,这表明马口鱼在游泳过程中更早且更多的依靠无氧代谢供能。已有研究报道,在鱼类中由于种类的不同和生境的差异常常会导致无氧代谢供能比例的差异[35]。本研究中,两种生境不完全重叠的实验鱼也存在着无氧代谢供能比例的差异。另外在本研究中,两种实验鱼的MO2active也随着溶氧水平的降低而下降,并且两中实验鱼的MO2active随溶氧水平下降的表现不一致,马口鱼的MO2active下降程度大于宽鳍鱲 (图 6);但马口鱼与宽鳍鱲的游泳速度下降比例并无显著差异,表明马口鱼在低氧下运动时有氧代谢供能不能满足其有氧运动的需求。由于马口鱼的低氧耐受能力强于宽鳍鱲,因此在低氧条件下马口鱼更多的利用无氧代谢为其有氧运动提供能量作为代谢补偿。 野外生存的两种生境不完全重叠的实验鱼不仅在低氧耐受能力方面存在显著差异,而且两者在不同低氧环境下的游泳运动能力及其运动过程中的代谢特征亦有不同表现,这很有可能是由于它们遗传特征、生存环境及生态习性差异所导致的结果。
| [1] | Chapman L J, Chapman C A, Nordlie F G, Nordlie F G, Rosenberger A E. Physiological refugia: swamps, hypoxia tolerance and maintenance of fish diversity in the Lake Victoria region. Comparative Biochemistry and Physiology Part A: Molecular and Integrative Physiology, 2002, 133(3): 421-437. |
| [2] | Fu S J, Peng Z G, Cao Z D, Peng J L, He X K, Xu D D, Zhang A J, Halsey L G. Habitat-specific locomotor variation among Chinese hook snout carp (Opsariichthys bidens) along a river. PLOS ONE, 2012, 7(7): e40791. |
| [3] | Diaz R J, Breitburg D L. The hypoxic environment. Fish Physiology, 2009, 27: 1-23. |
| [4] | Ultsch G R, Boschung H, Ross M J. Metabolism, critical oxygen tension, and habitat selection in darters (Etheostoma). Ecology, 1978, 59(1): 99-107. |
| [5] | Mandic M, Lau G Y, Nijjar M S, Richards J G. Metabolic recovery in goldfish: a comparison of recovery from severe hypoxia exposure and exhaustive exercise. Comparative Biochemistry and Physiology-Part C: Toxicology and Pharmacology, 2008, 148(4): 332-338. |
| [6] | Shingles A, Mckenzie D A, Claireaux G, Domenici P. Reflex cardioventilatory responses to hypoxia in the flathead gray Mullet (Mugilcephalus) and their behavioral modulation by perceived threat of predation and water turbidity. Physiological and Biochemical Zoology, 2005, 78(5): 744-755. |
| [7] | Wagner E J, Arndt R E, Brough M. Comparative tolerance of four stocks of cutthroat trout to extremes in temperature, salinity and hypoxia. Western North American Naturalist, 2001, 61(4): 434-444. |
| [8] | Waller U. Respiration and low oxygen tolerance of two fish species from the Arabian Sea, Cubiceps whiteleggi and Coryphaena hippurus. Journal of Applied Ichthyology, 1989, 5(3): 141-150. |
| [9] | Brauner C J, Matey V, Wilson J M, Bernier N J, Val A L. Transition in organ function during the evolution of air-breathing; insights from Arapaima gigas, an obligate air-breathing teleost from the Amazon. The Journal of Experimental Biology, 2004, 207(9): 1433-1438. |
| [10] | Petersen L H, Gamperl A K. Cod (Gadus morhua) cardiorespiratory physiology and hypoxia tolerance following acclimation to low-oxygen conditions. Physiological and Biochemical Zoology, 2011, 88(1): 18-31. |
| [11] | Stierhoff K L, Tyler R M, Targett T E. Hypoxia tolerance of juvenile Weakfish (Cynoscion regalis): laboratory assessment of growth and behavioral avoidance responses. Journal of Experimental Marine Biology and Ecology, 2009, 381(1): S173-S179. |
| [12] | Reidy S P, Kerr S R, Nelson J A. Aerobic and anaerobic swimming performance of individual Atlantic God. The Journal of Experimental Biology, 2000, 203(2): 347-357. |
| [13] | Plaut I. Critical swimming speed: its ecological relevance. Comparative Biochemistry and Physiology-Part A: Molecular and Integrative Physiology, 2001, 131(1): 41-50. |
| [14] | Zhang W, Cao Z D, Peng J L, Chen B J, Fu S J. The effects of dissolved oxygen level on the metabolic interaction between digestion and locomotion in juvenile southern catfish (Silurus meridionalis Chen). Comparative Biochemistry and Physiology-Part A: Molecular and Integrative Physiology, 2010, 157(3): 212-219. |
| [15] | Zhang W, Cao Z D, Fu S J. The effects of dissolved oxygen levels on the metabolic interaction between digestion and locomotion in Cyprinid fishes with different locomotive and digestive performances. Journal of Comparative Physiology B: Biochemical, Systemic and Environmental Physiology, 2012, 182(5): 641-650. |
| [16] | Fu S J, Brauner C J, Cao Z D, Richards J G, Peng J L, Dhillon R, Wang Y X. The effect of acclimation to hypoxia and sustained exercise on subsequent hypoxia tolerance and swimming performance in goldfish (Carassius auratus). The Journal of Experimental Biology, 2011, 214(12): 2080-2088. |
| [17] | Yeager D P, Ultsch G R. Physiological regulation and conformation: a basic program for the determination of critical points. Physiological Zoology, 1989, 62(4): 888-907. |
| [18] | Claireaux G, Couturier C, Groison A L. Effect of temperature on maximum swimming speed and cost of transport in juvenile European sea bass (Dicentrarchus labrax). The Journal of Experimental Biology, 2006, 209(17): 3420-3428. |
| [19] | Lee C G, Farrell A P, Lotto A, Hinch S G, Healey M C.. Excess post-exercise oxygen consumption in adult sockeye (Oncorhynchus nerka) and coho (O. kisutch) salmon following critical speed swimming. The Journal of Experimental Biology, 2003, 206(18): 3253-3260. |
| [20] | Zhang W, Cao Z D, Fu S J. Effect of dissolved oxygen level on metabolic mode in juvenile crucian carp. Acta Ecologica Sinica, 2012, 32(18): 5806-5812. |
| [21] | Scott G R, Wood C M, Slomand K A, Iftikar F I, De Boeck G, Almeida-Val V M F, Val A L.. Respiratory responses to progressive hypoxia in the Amazonian oscar, Astronotus ocellatus. Respiratory Physiology and Neurobiology, 2008, 162(2): 109-116. |
| [22] | Muusze B, Marcon J, Van den Thillart G, Almeida-Val V. Hypoxia tolerance of Amazon fish: Respirometry and energy metabolism of the cichlid Astronotus ocellatus. Comparative Biochemistry and Physiology-Part A: Molecular and Integrative Physiology, 1998, 120(1): 151-156. |
| [23] | Almeida-Val V M F, Val A L, Duncan W P, Souza F C A, Paula-Silva M N, Land S. Scaling effects on hypoxia tolerance in the Amazon fish Astronotus ocellatus (Perciformes: Cichlidae): contribution of tissue enzyme levels. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 2000, 125(2): 219-226. |
| [24] | Chapman L J, Kaufman L S, Chapman C A. Hypoxia tolerance in twelve species of east African cichlids: potential for low oxygen refugia in lake Victoria. Conservation Biology, 1995, 9(5): 1274-1287. |
| [25] | Graham J B, Dewar H, Lai N C, Lowell W R, Arce S M. Aspects of shark swimming performance determined using a large water tunnel. The Journal of Experiment Biology, 2004, 151(1): 175-192. |
| [26] | Li K G, Cao Z D, Fu S J. Effect of feeding on excess post-exercise oxygen consumption in juvenile Chinese catfish (Silurus asotus Linnaeus). Acta Hydrobiologica Sinica, 2012, 36(6): 1035-1040. |
| [27] | Gamperl A K, Rodnick K J, Faust H A, Venn E C, Bennett M T, Crawshaw L I, Keeley E R, Powell M S, Li H W..Metabolism, swimming performance, and tissue biochemistry of high desert redband Trout (Oncorhynchus mykiss ssp.): evidence for phenotypic differences in physiological function. Physiological and Biochemical Zoology, 2002, 75(5): 413-431. |
| [28] | Fu S J, Pang X, Cao Z D, Peng J L, Yan G J. The effects of fasting on the metabolic interaction between digestion and locomotion in juvenile southern catfish (Silurus meridionalis). Comparative Biochemistry and Physiology-Part A: Molecular and Integrative Physiology, 2011, 158(4): 498-505. |
| [29] | Fu S J, Zeng L Q, Li X M, Pang X, Cao Z D, Peng J L, Wang Y X.. The behavioural, digestive and metabolic characteristics of fishes with different foraging strategies. The Journal of Experimental Biology, 2009, 212(14): 2296-2302. |
| [30] | Nelson J A, Gotwalt P S, Snodgrass J W. Swimming performance of blacknose dace (Rhinichthys atratulus) mirrors home-stream current velocity. Canadian Journal of Fisheries and Aquatic Sciences, 2003, 60(3): 301-308. |
| [31] | Fitzgibbon Q P, Srawbridge A, Seymour R S. Metabolic scope, swimming performance and the effects of hypoxia in the mulloway, Argyrosomus japonicus (Pisces: Sciaenidae). Aquaculture, 2007, 270(1/4): 358-368. |
| [32] | Day N, Butler P J. The effects of acclimation to reversed seasonal temperatures on the swimming performance of adult brown trout Salmo trutta. The Journal of Experimental Biology, 2005, 208(18): 2683-2692. |
| [33] | Rome L C, Loughna P T, Goldspink G. Muscle fiber activity in carp as a function of swimming speed and muscle temperature. American Journal of Physiology, 1984, 247(2): R272-R279. |
| [34] | Burgetz I J, Rojas-vargas A, Hinch S G, Randall D J. Initial recruitment of anaerobic metabolism during submaximal swimming in rainbow trout (Oncorhynchus mykiss). The Journal of Experimental Biology, 1998, 201: 2711-2721 |
| [35] | Farrell A P. Cardiorespiratory performance during prolonged swimming tests with salmonids: a perspective on temperature effects and potential analytical pitfalls. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 2007, 362(1487): 2017-2030. |
| [20] | 张伟, 曹振东, 付世建. 溶氧水平对鲫鱼代谢模式的影响. 生态学报, 2012, 32(18): 5806-5812. |
| [26] | 李可贵, 曹振东, 付世建. 摄食对鲇鱼幼鱼力竭性运动后过量耗氧的影响. 水生生物学报, 2012, 36(6): 1035-1040. |