 2014, Vol. 34
2014, Vol. 34文章信息
- 李超, 彭赫, 程登发, 郭文超, 刘怀, 张云慧, 孙京瑞
- LI Chao, PENG He, CHENG Dengfa, GUO Wenchao, LIU Huai, ZHANG Yunhui, SUN Jingrui
- 马铃薯甲虫成虫田间扩散规律
- Diffusion of colorado potato beetle, Leptinotarsa decemlineata, adults in field
- 生态学报, 2014, 34(2): 359-366
- Acta Ecologica Sinica, 2014, 34(2): 359-366
- http://dx.doi.org/10.5846/stxb201304180743
-
文章历史
- 收稿日期:2013-4-18
- 修订日期:2013-9-22






2. 中国农业科学院植物保护研究所, 植物病虫害生物学国家重点实验室, 北京 100193;
3. 新疆农业科学院植物保护研究所, 乌鲁木齐 830000
2. State Key Laboratory for Biology of Plant Diseases and Insect Pests, Institute of Plant Protection Chinese Academy of Agricultural Sciences, Beijing 100193, China;
3. Institute of Plant Protection, Xinjiang Academy of Agricultural Sciences, Urumqi 830000, China
马铃薯甲虫是国际公认危害马铃薯的毁灭性害虫,也是我国重大外来入侵物种和一类植物检疫对象[1, 2, 3],自主扩散能力很强[4, 5]。在过去的100多年里,马铃薯甲虫从美国落基山东部地区逐步传播扩散到北美洲、欧洲、亚洲、非洲等40多个国家[6, 7, 8]。马铃薯甲虫以幼虫和成虫危害马铃薯等茄科作物,其危害通常是毁灭性的,每年给世界马铃薯生产造成十分严重的损失。马铃薯甲虫的扩散行为多发生在越冬滞育前后,在我国新疆北部地区多见于秋季和翌年初夏时节[9]。对于马铃薯甲虫在农田生态系统下的分布扩散规律[10],国外学者已开展了相关研究。美国学者运用“标记—释放—回捕”的方法[11]对马铃薯甲虫在田间扩散情况进行了研究[12]。结果表明释放的马铃薯甲虫更趋向于沿着释放时所在的垄上进行扩散,而很少进行跨行迁移。马铃薯甲虫自1993年入侵我国新疆北部地区以来,有关马铃薯甲虫在室内条件下的迁飞能力的研究的报道较多[13, 14, 15, 16],但在田间自然条件下马铃薯甲虫的扩散特性方面尚未见报道。本研究利用标记的方法对马铃薯甲虫不同世代成虫在马铃薯田块内的扩散规律展开研究,由于马铃薯甲虫在寄主田块内的扩散为短距离扩散,一般通过爬行的方式[1, 17, 18],本论文重点马铃薯甲虫在马铃薯田块内这一小范围内的扩散规律,不考虑通过飞行的方式进行的长距离扩散规律,旨在明确马铃薯甲虫在农田单一作物生态系统下的扩散特性,为后续研究更加复杂多样的生态环境下其扩散规律奠定基础。
1 材料与方法 1.1 试验虫源准备供试昆虫均采自新疆农科院综合试验场内的马铃薯田块,越冬代成虫采集于6月上旬,第1代成虫和第2代成虫则分别选择第1代和第2代老熟的四龄幼虫,让其在室内条件下羽化,第1代老熟四龄幼虫采集于7月上旬,第2代幼虫采集于8月上旬。释放时间为越冬代成虫,6月30日释放开始试验;第1代成虫,7月16日释放进行试验;第2代成虫,8月2日释放进行试验,试验开始前将成虫饲养48 h,待其鞘翅坚硬后,再进行标记。每次释放的虫口数为600头。
1.2 试验地概况试验地位于新疆维吾尔自治区乌鲁木齐县安宁渠镇(43°57′0.06″N,87°28′16.33″E),该田块连年种植马铃薯,是马铃薯甲虫越冬寄主田,面积约0.6 hm2。马铃薯品种为紫花白,播种时间为2012年4月30日,周边种植作物有玉米、豇豆、以及长年种植的榆树林,试验田北部为试验场内道理,宽约5 m,道路与豇豆田块之间有一行杨树作为防风林,以及水渠隔开,马铃薯田块内的株距、行距分别为0.25 m×0.75 m,种植垄呈东西排列。
1.3 试验方法试验在2012年夏季(6—8月)进行,将供试的昆虫准备好后用白色修正液在其鞘翅上做好标记,雌虫标记鞘翅的右边部分,雄虫标记左边部分,以便在田间释放后观察其扩散情况。在试验田块中心点来释放标记过的马铃薯甲虫成虫。释放后1,3,5,7,15 d定时(18:00—20:00)调查标记马铃薯甲虫在试验田内的分布情况。
调查时对试验田内进行全面系统的调查,即从释放点开始,沿马铃薯种植行依次进行,每隔1 m记录一次标记成虫的数量,马铃薯种植垄上1 m范围内所有马铃薯植株上的标记成虫全部统计。
1.4 数据处理与分析 1.4.1 扩散速度马铃薯甲虫成虫个体的扩散速度用该个体与释放点的直线距离除以其到达该点所耗时间得到。
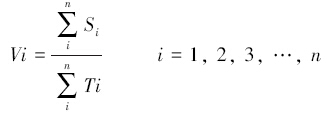
式中,Si为扩散后成虫的位置;Ti为调查时间。
1.4.2 聚集指标聚集度指标(m*/m):Lloyd定义平均拥挤度(m*)与平均密度(m)的比值为拥挤度指标。平均拥挤度指标表示平均每个个体与多少个其他个体在同一样方中。按照Lloyd的判断法则:当m*/m=1时,种群为随机分布;当m*/m<1时,种群为均匀分布;当m*/m>1时,种群为聚集分布[19]:
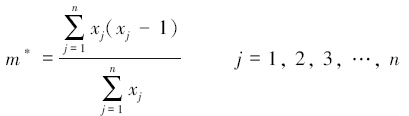
式中,m*为平均拥挤度;m为平均密度;xj为同一样方内的个体数量。
不同世代之间马铃薯甲虫的扩散速度的差异运用单因素方差分析(One-way ANOVA)进行比较,不同性别之间马铃薯甲虫扩散速度采用t-test检验分析,而对应各世代中不同扩散方向上的回捕虫量和扩散速度,由于得到的取样为关联样本,因此采用非参数Kruskal-Wallis检验进行比较,所有数据采用SPSS16.0软件进行处理。
2 结果与分析 2.1 不同世代马铃薯甲虫扩散速度分析不同世代的马铃薯甲虫成虫释放后,回捕得到虫口数量均随时间推移逐渐减少,回捕率逐渐下降。具体分析如下:释放1 d后,越冬代马铃薯田成虫回捕得到(277.3±4.06)头,第1代(272.3±4.33)头,第2代(235.7±4.98)头,回捕率分别为(46.22±0.68)%,(45.39±0.72)%,(39.28±0.83)%,且差异极显著(F=25.88,df=2,P<0.01);释放3 d后,越冬代马铃薯田成虫回捕得到(225±2.83)头,第1代(148±3.46)头,第2代(100.7±2.60)头,回捕率分别为(37.5±0.48)%,(24.7±0.58)%,(16.8±0.43)%,且差异极显著(F= 435.77,df=2,P<0.001);释放5 d后,越冬代马铃薯田成虫回捕得到(116±2.31)头,第1代(87±2.32)头,第2代(50±3.79)头,回捕率分别为(19.3±0.38)%,(14.5±0.38)%,(8.3±0.63)%,且差异极显著(F=131.32,df=2,P<0.001);释放7 d后,越冬代马铃薯田成虫回捕得到(66±2.31)头,第1代(53.3±2.60)头,第2代(35±1.73)头,回捕率分别为(11±0.38)%,(8.9±0.43)%,8(.3±0.29)%,且差异极显著(F= 48.23,df=2,P<0.001);释放15 d后,各世代回捕得到的标记成虫数量几乎为零。

|
| 图 1 试验地概况及马铃薯种植情况 Fig.1 Experimental field and potato planted pattern |

|
| 图 2 释放后不同时间后不同世代马铃薯甲虫回捕率分析 (新疆,2012) Fig.2 The recapture rate of different generations of CPB after releasing (Xinjiang,2012) |
不同世代马铃薯甲虫成虫扩散速度差异显著(F=4.45,df=2,P<0.05)越冬代成虫和第2代成虫扩散速度显著高于第1代成虫,而越冬代和第2代成虫之间比较,两者扩散速度差异不明显,分别为2.64,2.52 m/d。马铃薯甲虫释放后,随着时间的推移,其扩散速度也在发生变化。越冬代成虫在释放前期(1 d和3 d)扩散速度较低,中期(5 d)扩散速度达到最大,3.44 m/d,随后扩散速度逐渐下降。第1代成虫扩散速度随时间波动变化较小,扩散速度多维持在1.66 m/d。第2代成虫则在释放后前期(1 d)扩散速度最高,达3.51 m/d,后随时间推移逐渐降低,但在释放后期(15 d),其扩散速度有所上升,达到2.59 m/d(图 3)。

|
| 图 3 不同世代马铃薯甲虫释放后不同时间内扩散速度 (新疆,2012) Fig.3 The curves of diffusion velocity of different generations after release (Xinjiang,2012) |
结果如图 4所示,试验田内调查时,不同调查时间回捕得到的马铃薯甲虫雌虫数量均多于雄虫数量。在扩散速度上(图 5),雌虫扩散速度为(2.74±0.25) m/d,雄虫扩散速度仅为雌虫一半,即(1.33±0.28) m/d。释放后不同时间内,雌虫扩散速度均大于雄虫,且差异极显著(F=0.143,df=8,P<0.01)。同时,释放不同时间后,马铃薯甲虫雌虫和雄虫的扩散速度变化趋势,基本一致,其中雄虫扩散速度跟随雄虫扩散速度变化。

|
| 图 4 不同性别的马铃薯甲虫成虫的回捕数量 (新疆,2012) Fig.4 The insect amount of recapture of different genders after release (Xinjiang,2012) |

|
| 图 5 不同性别的马铃薯甲虫成虫扩散速度 (新疆,2012) Fig.5 The diffusion velocity of different genders (Xinjiang,2012) |
越冬代马铃薯甲虫对扩散方向的选择如图 6所示,对不同方向上回捕得到的马铃薯甲虫虫口数量和扩散速度进行非参数检验差异显著,结果显示,不同方向上回捕得到的虫量差异不显著,不同方向上的马铃薯甲虫成虫的扩散速度差异也不显著。

|
| 图 6 马铃薯甲虫成虫在不同方向上的回捕虫量和扩散速率的比较分析 (新疆,2012) Fig.6 The recapture amount and diffusion velocity of CPB in different directions after release (Xinjiang,2012) 不同大写字母表示第1代成虫在不同扩散方向上回捕虫量与扩散速率的差异(P<0.05,非参数检验 |
释放不同时间后,各方向上回捕得到的虫量分析可知,在西北方向上的回捕虫量均为最多,1 d后,回捕得到122头;释放3 d后,回捕得到121头;释放5 d后,回捕得到46头;释放后7 d后,回捕得到32头。 同时,马铃薯甲虫成虫在田间的平均拥挤度、平均密度和拥挤度指标均随释放后时间的增大而逐渐减小,至释放后7 d,马铃薯甲虫的空间分布从聚集分布逐渐变化为均匀分布(表 1)。而在不同扩散方向上,越冬代成虫的扩散速度无明显规律。
|
世代
Generations | 释放后时间/d
Time after release | 平均拥挤度
Mean crowding(m*) | 平均密度
Mean density(m) | 拥挤度指标
Aggregation index(m*/m) |
| 越冬代Overwintering | 1 | 24.749 | 5.642 | 4.387 |
| 3 | 4.221 | 2.036 | 2.073 | |
| 5 | 0.500 | 1.137 | 0.440 | |
| 7 | 0.294 | 1.115 | 0.264 | |
| 第1代First summer | 1 | 38.347 | 6.949 | 5.519 |
| 3 | 4.828 | 2.042 | 2.364 | |
| 5 | 0.927 | 1.390 | 0.667 | |
| 7 | 0.087 | 1.045 | 0.083 | |
| 第2代Second summer | 1 | 17.670 | 4.816 | 3.669 |
| 3 | 1.248 | 1.485 | 0.840 | |
| 5 | 0.078 | 1.041 | 0.075 | |
| 7 | 0 | 1 | 0 |
根据非参数检验分析结果,第1代马铃薯甲虫成虫在不同方向上回捕的虫量差异显著(U=8.72,df=3,P<0.05),在不同方向上扩散速度差异也显著(U=7.92,df=3,P<0.05)。 释放不同时间后,各方向上回捕得到的虫量分析可知,除释放7 d后回捕得到的马铃薯甲虫成虫数量在西南方向上最多外,其他时间在西北方向上的回捕虫量均为最多:1 d后,回捕得到149头;释放3 d后,回捕得到85头;释放5 d后,回捕得到38头。释放后7 d后,西北方向回捕得到15头,而西南方向上得到17头。同时,马铃薯甲虫成虫在田间的平均拥挤度、平均密度和拥挤度指标均随释放后时间的增大而逐渐减小,至释放后7 d,马铃薯甲虫的空间分布从聚集分布逐渐变化为均匀分布(表 1)。而在不同扩散方向上,第1代成虫的扩散速度无明显规律。
2.3.3 第2代成虫扩散方向运用非参数检验比较第2代成虫在不同方向上的回捕虫量和扩散速度,结果表明,不同方向上回捕虫量差异不显著,不同方向上的扩散速度差异不显著。释放1 d后,在西北方向上回捕虫量最多,为125头,释放3 d后,回捕虫量最多的方向为西南和东南方向上,虫量均为27头,释放5 d后,东南方向上回捕虫量最多,为20头,释放后7 d后,东北方向上回捕得到的虫量最多,均为12头。随着释放后时间的延长,第2代马铃薯甲虫成虫不同方向上回捕得到的虫量逐渐减少。同时,马铃薯甲虫成虫在田间的平均拥挤度、平均密度和拥挤度指标均随释放后时间的增大而逐渐减小,至释放后7 d,马铃薯甲虫的空间分布从聚集分布逐渐变化为均匀分布(表 1)。而在不同扩散方向上,第2代成虫的扩散速度无明显规律。
3 结论与讨论爬行是马铃薯甲虫成虫在田间扩散的主要方式[20]。田间寄主植物的匮乏或低劣的食物质量是导致马铃薯甲虫大量扩散的原因[21],由于越冬代马铃薯甲虫成虫出土后,食物的匮乏会迫使其寻找寄主植物,补充营养以完成生活史,而第2代马铃薯甲虫成虫的发生高峰期在8月中下旬至9月上旬,此时早熟的马铃薯品种已经收获,晚熟的品种也已转入成熟期,植株长势衰弱给马铃薯甲虫的食物供给带来问题,扩散转移以获得新的食物来源是马铃薯甲虫的必然选择。而相比越冬代与第2代成虫,第1代成虫的发生时期食物来源充沛。由此可以推测寄主植物的匮乏也是导致3种世代的马铃薯甲虫扩散能力差异的关键因素。同时,国内学者在室内条件下对马铃薯甲虫飞行能力的测试的结果也表明短时间的营养缺乏容易引起马铃薯甲虫的扩散飞行[13],同时虫口密度增大也在一定程度上造成了食物的匮乏,从而影响马铃薯甲虫的扩散[16]。越冬后饥饿的马铃薯甲虫飞行距离大于取食的马铃薯甲虫[8, 22]。
本研究着眼于农田生态系统下的马铃薯甲虫扩散规律,分别对3种世代成虫的扩散特点进行研究。随着时间的推移,马铃薯甲虫迁出调查田块的数量逐步增加,到释放15 d时,被标记的马铃薯甲虫的几乎全部迁出田外。由于马铃薯甲虫在实验田内多采用爬行的方式进行扩散,在实验田内回捕得到的虫口数量多,则表明选择爬行方式扩散的马铃薯甲虫多,其余未调查到的标记马铃薯甲虫成虫大部分则可能通过飞行的方式扩散到周边田块甚至更远处(排除少部分死亡个体)。郭丽娜等在室内条件下运用飞行磨对不同温度条件下马铃薯甲虫的飞行能力进行了研究,结果表明雄虫飞行能力稍大于雌虫,但无显著性差异[15]。与本文研究结果得到雌虫在田间的扩散能力显著强于雄虫这一结论存在一定差异。
本研究结果得到的马铃薯甲虫扩散速度只是一个低限的估测,因为马铃薯甲虫的扩散行为一般不会是连续地直线移动。在其扩散过程中可能会不断地改变运动方向,甚至可能会来回折返移动。因此马铃薯甲虫实际的迁移距离会大于通过释放——重捕来估测的扩散距离。同时本研究中,对马铃薯甲虫个体扩散距离的估测采用的是回捕点与释放点的直线距离,马铃薯田内沟垄交替,以及马铃薯植株自身的高度造成了马铃薯甲虫成虫实际扩散距离比直线距离要大得多。故本研究估测的速度扩散速度也是小于实际的扩散速度的。此外,本试验采用标记的马铃薯甲虫,也可能比未标记的正常马铃薯甲虫的扩散距离要短些。因为昆虫在标记过程中可能对其造成一定损伤,扩散能力有所降低。
明确马铃薯甲虫在田间的扩散规律,为马铃薯甲虫的田间监测提供了依据[23]。田间马铃薯甲虫的扩散受其栖息环境的影响,作物的覆盖作用对马铃薯甲虫的田间迁移起到了一定的阻碍作用。由于在650 m范围内马铃薯甲虫成虫的回捕率在30%左右,为有效的降低马铃薯甲虫越冬虫口基数,实施轮作的有效距离至少≥0.5 km[11]。同时,马铃薯甲虫的取食活动会受到多重景观结构的影响,为马铃薯甲虫的综合防控提供了新的思路:增加寄主密度和多样性能减小各阶段马铃薯甲虫的数量[24],非寄主田的存在能降低其定殖成功[25, 26]。本研究后续将开展马铃薯甲虫在多种作物混合种植等复杂生境下的扩散规律,为实际生产中马铃薯甲虫的监测防控提供理论依据。
| [1] | Alyokhin A. Colorado potato beetle management on potatoes: current challenges and future prospects// Tennant P, Benkeblia N, eds. Fruit, Vegetable and Cereal Science and Biotechology. Isleworth: GSB Publisher, 2009, 3: 10-19. |
| [2] | Alyokhin A, Baker M, Mota-Sanchez D, Dively G, Grafius E. Colorado potato beetle resistance to insecticides. American Journal of Potato Research, 2008, 85(6): 395-413. |
| [3] | Zhang R Z. Study of occurrence pattern and control techniques of Colorado potato beetle. Inistitute of Zoology, Chinese Academy of Sciences, Beijing, 1997. |
| [4] | Casagrande R A. The Iowa potato beetle, its discovery and spread to potatoes. Bulletin of the Entomological Society of America, 1985, 31 (2): 27-29. |
| [5] | Casagrande R A. The Colorado potato beetle: 125 years of mismanagement. Bulletin of the Entomological Society of America, 1987, 33 (3): 142-150. |
| [6] | Hare J D. Ecology and management of the Colorado potato beetle. Annual Review of Entomology, 1990, 35 (1): 81-100. |
| [7] | Caprio M A. Flight initiation behavior and host plant attraction in the Colorado potato beetle, Leptinotarsa decemlineata (Say) (Coleoptera: Chrysomelidae). Michigan State University. Department of Entomology, 1987. |
| [8] | Caprio M A, Grafius E J. Effects of light, temperature, and feeding status on flight initiation in postdiapause Colorado potato beetles (Coleoptera: Chrysomelidae). Environmental Entomology, 1990, 19 (2): 281-285. |
| [9] | Ahemaiti T, Xu J J, Guo W C, Liu J, He J, Xia Z H, Fu W J, Zhang D M. Study on major biological characteristics and occurrence regulation of Colorado potato beetle. Xinjiang Agricultural Sciences, 2010, 47(6): 1147-1151. |
| [10] | Alvarez J M, Srinivasan, R, Cervantes, F A. Occurrence of the Carabid beetle, pterostichus melanarius (Illiger), in potato ecosystems of Idaho and its predatory potential on the Colorado potato beetle and aphids. American Journal of Potato Research, 2012: 1-10. |
| [11] | Follett P A, Cantelo W W, Roderick G K. Local dispersal of overwintered Colorado potato beetle (Chrysomelidae: Coleoptera) determined by mark and recapture. Environmental Entomology, 1996, 25 (6): 1304-1311. |
| [12] | Szendrei Z, Grafius E, Byrne A, Ziegler A. Resistance to neonicotinoid insecticides in field populations of the Colorado potato beetle (Coleoptera: Chrysomelidae). Pest Management Science, 2012, 68 (6): 941-946. |
| [13] | Guo L N, Guo W C, Ahemaiti T, He J, Xu J J. Effects of different nutrients on flight capacity in Colorado potato beetles. Xinjiang Agricultural Sciences, 2012, 49 (3): 461-469. |
| [14] | Guo L N, Guo W C, Ahemaiti T, He J, Xu J J. Effects of host plants on flight capacity in Colorado potato beetles. Xinjiang Agricultural Sciences, 2011, 48 (5): 853-858. |
| [15] | Guo L N, Guo W C, Ahemaiti T, He J, Xu J J. Effect of temperature and feeding status on flight capacity of overwintering Colorado potato beetle. Plant Protection, 2011, 37 (5): 56-61. |
| [16] | Guo L N, Guo W C, Liu M S, Ahemaiti T, He J, Xu J J. Effects of density on flight capacity of Colorado potato beetles. Xinjiang Agricultural Sciences, 2011, 48 (2): 320-327. |
| [17] | Voss R H, Ferro D N. Ecology of migrating Colorado potato beetles (Coleoptera: Chrysomelidae) in western Massachusetts. Environmental Entomology, 1990, 19 (1): 123-129. |
| [18] | Voss R H, Ferro D N. Phenology of flight and walking by Colorado potato beetle (Coleoptera: Chrysomelidae) adults in western Massachusetts. Environmental Entomology, 1990, 19 (1): 117-122. |
| [19] | Xu R M, Cheng X Y. Basics and frontiers in population ecology of insects. Beijing: Science Press, 2005. |
| [20] | Boiteau G. Recruitment by flight and walking in a one-generation Colorado potato beetle (Coleoptera: Chrysomelidae) environment. Environmental Entomology, 2001, 30 (2): 306-317. |
| [21] | Sandeson P, Boiteau G, Le Blanc J R. Adult density and the rate of Colorado potato beetle (Coleoptera: Chrysomelidae) flight take-off. Environmental Entomology, 2002, 31 (3): 533-537. |
| [22] | Ferro D N, Alyokhin A V, Tobin D B. Reproductive status and flight activity of the overwintered Colorado potato beetle. Entomologia Experimentalis et Applicata, 1999, 91 (3): 443-448. |
| [23] | Werling B P, Gratton C. Local and broadscale landscape structure differentially impact predation of two potato pests. Ecological Applications, 2010, 20 (4): 1114-1125. |
| [24] | Capinera J L, Horton D R, Epsky N D, Chapman P L. Effects of plant density and late-season defoliation on yield of field beans. Environmental Entomology, 1987, 16 (1): 274-280. |
| [25] | Bach C E. The influence of plant dispersion on movement patterns of the Colorado potato beetle, Leptinotarsa decemlineata(Coleoptera: Chrysomelidae). Great Lakes Entomologist, 1982, 15 (4): 247-252. |
| [26] | Zehnder G W, Hough G J. Colorado potato beetle (Coleoptera: Chrysomelidae) population development and effects on yield of potatoes with and without straw mulch. Journal of Economic Entomology, 1990, 83 (5): 1982-1987. |
| [3] | 张润志. 马铃薯甲虫发生规律和防治技术研究. 北京: 中国科学院动物所, 1997. |
| [9] | 吐尔逊·阿合买提, 许建军, 郭文超, 刘建, 何江, 夏正汉, 付文君, 张冬梅. 马铃薯甲虫主要生物学特性及发生规律研究. 新疆农业科学, 2010, 47(6): 1147-1151. |
| [13] | 郭文超, 吐尔逊, 郭利娜, 何江, 许建军. 营养对马铃薯甲虫迁飞能力的影响. 新疆农业科学, 2012, 49 (3): 461-469. |
| [14] | 郭利娜, 郭文超, 吐尔逊, 何江, 许建军. 寄主对马铃薯甲虫飞行能力的影响. 新疆农业科学, 2011, 48 (5): 853-858. |
| [15] | 郭利娜, 郭文超, 吐尔逊, 何江, 许建军. 温度与取食对越冬后马铃薯甲虫飞行能力的影响. 植物保护, 2011, 37 (5): 56-61. |
| [16] | 郭利娜, 郭文超, 刘曼双, 吐尔逊, 何江, 许建军. 拥挤度对马铃薯甲虫飞行能力的影响. 新疆农业科学, 2011, 48 (2): 320-327. |
| [19] | 徐汝梅, 成新跃. 昆虫种群动态学——基础与前沿. 北京:科学出版社, 2005. |