 2014, Vol. 34
2014, Vol. 34文章信息
- 孙鹏飞, 单秀娟, 吴强, 陈云龙, 金显仕
- SUN Pengfei, SHAN Xiujuan, WU Qiang, CHEN Yunlong, JIN Xianshi
- 莱州湾及黄河口水域鱼类群落结构的季节变化
- Seasonal variations in fish community structure in the Laizhou Bay and the Yellow River Estuary
- 生态学报, 2014, 34(2): 367-376
- Acta Ecologica Sinica, 2014, 34(2): 367-376
- http://dx.doi.org/10.5846/stxb201303060354
-
文章历史
- 收稿日期:2013-3-6
- 修订日期:2013-7-8






2. 山东省渔业资源与生态环境重点实验室 中国水产科学研究院黄海水产研究所, 青岛 266071
2. Shandong Provincial Key Laboratory of Fishery Resources and Ecological Environment (SFREE), Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Qingdao 266071, China
河口是生态环境十分脆弱和敏感的水域,许多理化特征和生物特征具有独特性[1],莱州湾及黄河口水域有黄河、小清河和弥河等多条河流入海,基础饵料丰富,是渤海的传统渔场,也是黄、渤海众多渔业生物的产卵场、育幼场和索饵场,对黄、渤海渔业资源的补充具有重要意义[1, 2]。从20世纪50年代到21世纪初,关于该水域渔业资源的研究广泛开展,主要包括资源结构、资源分布和多样性变化等[1, 3, 4, 5, 6, 7],然而近年来关于该水域鱼类群落结构的研究较少,仅见于李凡、王广成等的研究[8, 9],本文根据2011—2012年莱州湾及黄河口水域渔业底拖网调查数据,对该水域鱼类群落结构特征的季节变化进行了研究,以期为莱州湾及黄河口水域渔业生态系统结构和功能的变动及渔业资源的管理提供基础资料。
1 材料与方法 1.1 研究区域调查范围为莱州湾及黄河口(38°30′N以南,118°30′—120°30′E)水域,调查站位(18个)设置沿袭1982、1993和1998年3个时期在莱州湾及黄河口(38°30′N以南)进行的定点底拖网调查的站位,基本以10′×10′为原则尽可能覆盖调查对象的整个分布区域,剔除浅水区调查船无法进入的区域,并于河口区域适当添加站位(图 1)。
1.2 数据来源及处理数据取自2011年5—11月和2012年3—4月对调查区域的渔业底拖网调查,2011年5月、2012年3月和4月为春季,2011年6月、7月和8月为夏季,2011年9月、10月、11月为秋季,调查站位见图 1。调查船为“鲁昌渔4193”(350kW),平均拖速3kn,每站拖网1h。调查网具为单船底拖网,网具参数:网口周长30.6 m,网囊网目20mm,拖网时网口宽度约8m。渔获物中捕获的鱼类鉴定到种,记录每种鱼类的重量和尾数,平均单位时间渔获量以单位时间的渔获量(kg/h)和单位时间的渔获尾数(尾/h)表示。
1.3 研究方法 1.3.1 种类组成及优势种分析依据田明诚、邓景耀等对适温类型的分析方法[10, 11],将莱州湾及黄河口水域鱼类划分为暖水种、暖温种、冷温种和冷水种。

|
| 图 1 莱州湾及黄河口水域渔业调查站位 Fig.1 Sampling stations in the Laizhou Bay and Yellow River Estuary |
鱼类优势种由相对重要性指标(IRI)确定[12]:
IRI=(N+W)×F
式中,N为某种类的尾数占总渔获尾数的百分比; W为某种类的渔获量占总渔获量的百分比; F为某种类在调查中被捕获的站数与总调查站位百分比。
其中IRI值大于500为优势种,500—100为常见种,100—10为一般种,10—1为少见种,小于1为稀有种[13]。本文将优势种和常见种作为鱼类群落重要种类成分。
通过优势度曲线来分析各季节优势种的优势度变化[14]。
1.3.2 多样性指数用Margalef种类丰富度指数(R)、Shannon-Wiener多样性指数(H′)及Pielou均匀度指数(J′)分析莱州湾及黄河口水域鱼类多样性[15, 16, 17]。
Margalef种类丰富度指数R:
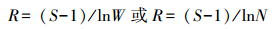
Shannon-Wiener多样度指数H′:
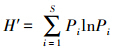
Pielou种类均匀度指数J′:
J′=H′/lnS
式中,S为种类数;W为总渔获量,N为总渔获尾数;Pi为第i种鱼所占总渔获量或总渔获尾数的比例。
另外,各站位多样性指数与环境因子的关系采用Pearson相关系数分析。
1.3.3 相似性指数用Bray-Curtis的相似性系数[18]计算各调查航次(季节)种类生物量及其组成的相似性,分析莱州湾及黄河口水域鱼类资源随时间变化的特征。
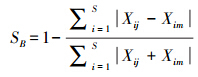
式中,SB为相似性系数,Xij和Xim分别为第i种在j和m航次调查中的渔获量或渔获尾数。
2 结果 2.1 种类组成2 011年5月至2012年4月莱州湾及黄河口水域共捕获鱼类62种,隶属于11目34科53属(表 1),其中鲈形目种类最多,有14科27属37种;其次为鲉形目,有6科7属7种;再次为鲽形目和鲱形目,鲽形目有4科5属6种,鲱形目有2科5属6种;其余各目均不超过4种。
| 目Order | 科Family | 属Genus | 种Species |
| 鲉形目 Scorpaeniformes | 6 | 7 | 7 |
| 银汉鱼目Atheriniformes | 1 | 1 | 1 |
| 鳐形目Rajiformes | 1 | 1 | 1 |
| 仙女鱼目Aulopiformes | 1 | 1 | 1 |
| 鲀形目Tetraodontiformes | 2 | 2 | 4 |
| 鲈形目Perciformes | 14 | 27 | 31 |
| 鲑形目Salmoniformes | 1 | 2 | 2 |
| 鲱形目Clupeiformes | 2 | 5 | 6 |
| 鲽形目Pleuronectiformes | 4 | 5 | 6 |
| 刺鱼目Gasterosteiformes | 1 | 1 | 2 |
| 鮟鱇目Lophiiformes | 1 | 1 | 1 |
春季优势种为矛尾鰕虎鱼、鲱衔、短吻红舌鳎、矛尾复鰕虎鱼和方氏云鳚,常见种为丝鰕虎鱼(Cryptocentrus filifer)、中华栉孔鰕虎鱼(Ctenotrypauchen chinensis)、细纹狮子鱼(Liparis tanakae)、 大银鱼(Protosalanx chinensis)和六丝矛尾鰕虎鱼(Amblychaeturichthys hexanema);夏季优势种为矛尾鰕虎鱼、鲱衔、斑鰶和短吻红舌鳎,常见种为鳀、蓝点马鲛(Scomberomorus niphonius)、小黄鱼、赤鼻棱鳀和矛尾复鰕虎鱼;秋季优势种为矛尾鰕虎鱼、赤鼻棱鳀、鳀、青鳞小沙丁鱼、斑鰶、小黄鱼和矛尾复鰕虎鱼,常见种为鲱衔、鲬鱼(Platycephalus indicus)和短吻红舌鳎(表 2)。
优势种渔获量在春、夏和秋季分别占总渔获量的70.8%、68.1%和87.1%,渔获尾数分别占58.1%、79.3%和95.4%。各季节矛尾鰕虎鱼均为第一优势种,其渔获量和渔获尾数在春、夏和秋季分别占总渔获量和总渔获尾数的38.8%、38.6%、18.2%和35.3%、55.5%、14.0%。春季鱼类重要种类成分中,底层鱼类9种,其渔获量占总渔获量的86.1%,中上层鱼类仅大银鱼一种,占总渔获量的1.4%;夏季重要种类成分中,底层鱼类5种,占总渔获量的57.3%,中上层鱼类4种,占总渔获量的31.7%;秋季重要种类成分中,底层鱼类6种,占总渔获量的41.3%,中上层鱼类4种,占总渔获量的50.4%。
| 季节Season | 种名Species | W/% | N/% | IRI |
| W,N分别表示各季节鱼类重量和尾数占总渔获重量和总渔获尾数的百分比 | ||||
| 春季Spring | 矛尾鰕虎鱼Chaeturichthys stigmatias | 38.8 | 35.3 | 6787 |
| 鲱衔Callionymus beniteguri | 6.8 | 9.8 | 1140 | |
| 短吻红舌鳎Cynoglossus joyneri | 4.4 | 8.2 | 948 | |
| 矛尾复鰕虎鱼Synechogobius hasta | 15.8 | 4.8 | 943 | |
| 方氏云鳚Enedrias fangi | 5.0 | 8.4 | 724 | |
| 丝鰕虎鱼Cryptocentrus filifer | 1.8 | 10.1 | 496 | |
| 中华栉孔鰕虎鱼Ctenotrypauchen chinensis | 1.3 | 8.5 | 487 | |
| 细纹狮子鱼Liparis tanakae | 11.3 | 0.2 | 215 | |
| 大银鱼Protosalanx chinensis | 1.4 | 5.9 | 182 | |
| 六丝矛尾鰕虎鱼Amblychaeturichthys hexanema | 1.0 | 3.5 | 131 | |
| 夏季Summer | 矛尾鰕虎鱼Chaeturichthys stigmatias | 38.6 | 55.5 | 8189 |
| 鲱衔Callionymus beniteguri | 6.0 | 9.7 | 1340 | |
| 斑鰶Konosirus punctatus | 17.6 | 11.9 | 1308 | |
| 短吻红舌鳎Cynoglossus joyneri | 5.9 | 2.2 | 628 | |
| 鳀Engraulis japonicus | 2.5 | 10.4 | 357 | |
| 蓝点马鲛Scomberomorus niphonius | 7.4 | 1.1 | 298 | |
| 小黄鱼Larimichthys polyactis | 3.0 | 0.30 | 231 | |
| 赤鼻棱鳀Thrissa kammalensis | 4.3 | 2.0 | 199 | |
| 矛尾复鰕虎鱼Synechogobius hasta | 3.0 | 2.0 | 139 | |
| 秋季Autumn | 矛尾鰕虎鱼Chaeturichthys stigmatias | 18.2 | 14.0 | 3005 |
| 赤鼻棱鳀Thrissa kammalensis | 9.6 | 19.6 | 1653 | |
| 鳀Engraulis japonicus | 17.7 | 42.3 | 1565 | |
| 青鳞小沙丁鱼Sardinella zunasi | 10.8 | 10.9 | 1372 | |
| 斑鰶Konosirus punctatus | 12.2 | 2.9 | 1179 | |
| 小黄鱼Larimichthys polyactis | 12.0 | 4.1 | 876 | |
| 矛尾复鰕虎鱼Synechogobius hasta | 6.6 | 1.6 | 551 | |
| 鲱衔Callionymus beniteguri | 1.4 | 1.9 | 269 | |
| 鲬鱼Platycephalus indicus | 2.0 | 0.2 | 155 | |
| 短吻红舌鳎Cynoglossus joyneri | 1.2 | 0.5 | 133 | |
莱州湾及黄河口水域暖水种、暖温种和冷温种鱼类分别有25种、28种和9种,分别占鱼类种类总数的40.3%、45.2%和14.5%。暖温种的平均单位时间渔获量和平均单位时间渔获尾数最多,暖水种次之,冷温种最少。暖水种类的平均单位时间渔获量和平均单位时间渔获尾数从春季至秋季呈上升趋势,由0.14kg/h和28尾/h上升至8.27kg/h和1757尾/h;暖温性种类平均单位时间渔获量从春季0.93kg/h上升至秋季14.16kg/h,夏季和秋季平均单位时间渔获尾数分别为3544尾/h和3260尾/h,春季仅为83尾/h;冷温性种类的平均单位时间渔获量和平均单位时间渔获尾数夏季最高,分别为0.70kg/h和108尾/h,春、秋季远低于夏季(图 2)。

|
| 图 2 鱼类平均网获量和平均网获尾数的季节变化 Fig.2 Seasonal variations of average weight and number of catch per haul |
春季暖温种渔获量最高,且多分布于近岸区域,暖水种渔获量最低,冷温种多分布于莱州湾东北部深水区域;夏季暖温种分布较广,暖水种多分布于莱州湾南部近岸区域;秋季暖水种和暖温种渔获量较高,多分布于莱州湾东北部深水区域(图 3)。
2.4 群落相似性、多样性及其影响因子鱼类种类数夏季最多,秋季次之,春季最少。从2011年5月至7月种类数增加,7月种类数最多(42种),此后至2012年4月一直下降,3月和4月仅为18种。
以重量或尾数计算的丰富度指数(R)均是夏季最大,春季最小,在1.650—3.317之间波动,呈先上升后下降趋势,其中以2011年6月最高,次年3月最低,11月小幅上升;以重量或尾数计算的多样性指数(H′)在1.480—2.410之间波动,夏季最高(2.023和1.979),秋季次之(1.603和1.622),春季最小,其中2011年5月最高,2012年3月最低;以重量计算的均匀度指数(J′)在0.411—0.716之间波动,春季最大(0.612),夏季最小(0.552),其中2011年5月最高,11月最低,以尾数计算的均匀度指数(J′)在0.388—0.670之间波动,其中以2011年5月最大,8月最小,夏季最大,春季最小(图 4)。

|
| 图 3 莱州湾及黄河口水域鱼类各适温类型渔获重量的分布 Fig.3 The distribution of the catches of each optimum temperature type of fish in the Laizhou Bay and Yellow River Estuary |

|
| 图 4 莱州湾及黄河口水域鱼类生物多样性指数的变化 Fig.4 Variations of fish species biodiversity indices in the Laizhou Bay and Yellow River Estuary Rw、Hw′、Jw′和 Rn、Hn′、Jn′分别表示以重量和数量计算的多样性指数 |
春、夏季鱼类优势度曲线比较接近,并且春、夏季优势种较秋季突出,春、夏和秋季最高单种分别占总渔获量的37.8%、38.6%和18.1%(图 5);夏季优势度曲线高于秋季,秋季高于春季,夏季优势种最突出,春、夏和秋季最高单种所占渔获尾数分别为35.3%、55.5%和42.3%(图 5)。
对鱼类种类丰富度指数(R)、多样性指数(H′)和均匀度指数(J′)与环境因子进行Pearson相关性分析(表 3),以重量或尾数计算的丰富度指数(R)与表层盐度呈极显著正相关(P<0.01),丰富度指数(R)和多样性指数(H′)与表层温度呈极显著相关(P<0.01),其中以尾数计算的多样性指数(H′)与之呈极显著负相关,以重量计算的种类均匀度指数(J′)与表层盐度呈极显著正相关(P<0.01),以重量计算则呈极显著负相关(P<0.01)。

|
| 图 5 莱州湾及黄河口水域鱼类优势度曲线 Fig.5 Dominance curves of fish in the Laizhou Bay and Yellow River Estuary |
| Pearson correlation | 盐度 Salinity | 温度 Temperature | 深度 Depth |
| Rw、Hw′、Jw′和 Rn、Hn′、Jn分别表示以重量和数量计算的多样性指数;**表示极显著相关 | |||
| Rw | 0.241** | 0.442** | 0.157 |
| Rn | 0.189** | 0.174** | 0.130 |
| Hw′ | 0.112 | 0.176** | 0.057 |
| Hn′ | 0.006 | -0.203** | 0.025 |
| Jw′ | 0.000** | -0.190 | -0.010 |
| Jn′ | -0.100** | -0.495 | -0.033 |
以重量或尾数计算的相似性趋势一致:相邻各月之间相似性较高;季节内各月之间相似性较高,夏季月份和秋季月份之间相似性较高,3月和11月、5月和11月之间的相似性也较高(表 4)。
| 年份 Year | 月份 Month | 2011 | 2012 | |||||||
| 5 | 6 | 7 | 8 | 9 | 10 | 11 | 3 | 4 | ||
| 对角线上方数字为以尾数计,下方数字为以重量计 | ||||||||||
| 2011 | 5 | 0.22 | 0.02 | 0.04 | 0.01 | 0.04 | 0.51 | 0.38 | 0.51 | |
| 2011 | 6 | 0.26 | 0.10 | 0.17 | 0.10 | 0.22 | 0.28 | 0.23 | 0.13 | |
| 2011 | 7 | 0.11 | 0.22 | 0.61 | 0.39 | 0.18 | 0.04 | 0.02 | 0.01 | |
| 2011 | 8 | 0.09 | 0.14 | 0.77 | 0.37 | 0.18 | 0.08 | 0.05 | 0.02 | |
| 2011 | 9 | 0.05 | 0.15 | 0.55 | 0.57 | 0.40 | 0.03 | 0.02 | 0.01 | |
| 2011 | 10 | 0.12 | 0.34 | 0.36 | 0.30 | 0.50 | 0.50 | 0.05 | 0.02 | |
| 2011 | 11 | 0.27 | 0.13 | 0.16 | 0.19 | 0.13 | 0.20 | 0.63 | 0.35 | |
| 2012 | 3 | 0.43 | 0.10 | 0.13 | 0.10 | 0.06 | 0.14 | 0.47 | 0.41 | |
| 2012 | 4 | 0.41 | 0.12 | 0.04 | 0.04 | 0.02 | 0.05 | 0.20 | 0.39 | |
莱州湾及黄河口水域是多种水生经济生物生长、发育的重要场所,鱼类是该区域生物群落的重要组成部分。本次调查捕获鱼类62种,仅为1982—1985年鱼类种类数的54.4%,1992—1993年捕获鱼类数的84.9%,若考虑调查网具差异,种类可能略有不同。从春季到秋季,鱼类群落重要种类成分中,底层鱼类在总渔获量中的比例由86.1%减少至41.3%,中上层鱼类在总渔获量中的比例由1.4%增加至50.4%,经济种类如短吻红舌鳎、鳀、赤鼻棱鳀等在渔获量中的比例增加。优势种由春季以底层经济种类为主,过渡到秋季以中上层小型经济种类为主,但是矛尾鰕虎鱼在3个季节均为第一优势种。自1950年代以来,莱州湾及黄河口水域的鱼类优势种发生了很大的变化,其中1959年以带鱼(Trichiurus lepturus)、小黄鱼为主;1982—1985年优势种为黄鲫(Setipinna taty)、黑鳃梅童鱼(Collichthys niveatus)、鳀和小黄鱼;1992—1993年鳀为绝对优势种;1998年赤鼻棱鳀和黄鲫占较大优势,鳀仅在春季为优势种,黄鲫在春、夏、秋季都为优势种[1, 3, 4, 5, 6],目前,莱州湾及黄河口水域渔业资源主要优势种群按黄鲫→鳀→赤鼻棱鳀顺序朝着更加小型化方向演替[1]。鱼类群落结构的这种演替可能与过度捕捞和环境退化使生物群落生态系统恢复力和完整性降低,生态系统的稳定性转差有关[19]。近年来选择性的商业过度捕捞使该水域经济鱼类迅速减少,超过其资源再生能力,造成资源严重衰退;由于大型经济鱼类减少,并且春、夏季洄游性鱼类到达该水域的数量较少[20],小型鱼类特别是底层鱼类被捕食的机会减少,使其数量迅速增加,成为该水域的优势种。另外,沿岸经济发展带来的陆源污染、黄河上游水利工程建设对径流量的调节、养殖工程建设造成的近岸栖息地物理环境的破坏都对渔业资源产生了重要的影响。如我国沿岸海域中,莱州湾海域鱼体内肌肉重金属镉的含量最高,重金属铅、汞、铬的含量均在前5位[21]。
3.2 渔获量及其分布暖温种和暖水种的平均单位时间渔获量和平均单位时间渔获尾数从春季到秋季逐渐升高,从秋季到翌年春季逐渐降低。这一方面与莱州湾及黄河口水域夏、秋季环境、温度等更适宜生物生长,并且夏、秋季饵料相对于冬、春季丰富有关[22];另一方面也与每年春季到秋季(4—11月)黄海鱼类从越冬场到渤海沿岸的产卵洄游有关[4],如小黄鱼6月开始在渤海各海湾、黄海北部沿岸和海州湾产卵,栖息在渤海的小黄鱼 9—11月在渤海中部索饵,11月后绕过成山头向越冬场洄游[23]。因此,春季由黄海游入莱州湾及黄河口水域的鱼类群体还比较少,秋季鱼类资源主要由补充群体组成,夏季饵料生物较为丰富,鱼类生长迅速,斑鰶、鳀、小黄鱼等优势度增加。考虑到调查网具差异,经过数据标准化[24],2011年春季(5月)平均单位时间渔获量为0.69kg/h,是1959年的0.3%,2003年的2.6%,2006年的1.7%,2008年的541.2%[25],莱州湾与黄河口水域鱼类资源量近年来虽有所回升,但总体呈下降的趋势,近50年来,该水域的捕捞努力量增加了40倍,高强度的捕捞努力量远超过渔业资源自身再生能力,渔业资源衰退,种类更替加快,优势种优势度下降,致使渔业资源结构发生明显变化,一些主要经济种类的甚至消失,难以维持渔业资源的可持续发展[26, 27, 28, 29, 30];海洋溢油、城市生活污水、沿岸养殖工程建设造成的环境污染日益严重,引起赤潮频发,且面积增大,破坏了原有生态系统结构,给渔业资源带来很大的负面影响[31, 32]。另外,黄河径流量的变化也对该水域理化环境和生物环境造成了一定影响,从而引起该水域的渔业资源结构和资源量的变化,如在黄河丰水期(6—8月)无机氮、无机磷和石油类的超标,并且磷限制、氮污染扩大,导致基础饵料生物的组成和资源量发生显著变化[33, 34, 35]。
3.3 多样性和相似性海洋鱼类群落结构由于物种洄游性季节分布导致其时序的相对不稳定性[5],多样性是表征群落结构和功能的重要因素[36]。莱州湾及黄河口水域夏、秋季鱼类种类数、丰富度指数和多样性指数较高,春季较低。春季多样性指数(0.20—2.50)高于1982年(0.70—2.07)、1993年(0.34—2.02)和1998年(0.50—1.99)对应月份的多样性指数[37],但要低于我国南海大陆架底层鱼类群落(1.40—4.10)和东海深海底层鱼类群落(0.25—4.40)的多样性[38, 39]。均匀度指数以春季最高,这与大型底层鱼类如细纹狮子鱼等数量较大密切相关[40]。鱼类丰富度指数(R)和多样性指数(H′)与表层温度呈极显著相关(P<0.01),丰富度指数(R)与表层盐度呈极显著正相关(P<0.01),均匀度指数(J′)与表层盐度呈极显著相关(P<0.01),夏、秋季平均表层温度和盐度较高,因此夏秋季多样性和相似性较高,春季较低。研究证明,多数自然群落处于非平衡状态,竞争性平衡被一些因子所阻止,如生物资源量周期性降低、生物死亡、物理环境和生物环境变动等[41]。当扰动为中等强度时多样性最高,当扰动处于两极时多样性较低[29, 36],目前,莱州湾及黄河口水域维持了相对高的生物多样性,处于中度干扰强度,但支持了相对较低的生物量,这在长江口水域鱼类群落结构中也有类似发现[42]。
| [1] | Jin X S, Deng J Y. Variations in community structure of fishery resources and biodiversity in the Laizhou Bay, Shandong. Chinese Biodiversity, 2000, 8(1): 65-72. |
| [2] | Deng J Y, Jin X S. Study on fishery biodiversity and its conservation in Laizhou Bay and Yellow River estuary. Zoological Research, 2000, 21(1): 76-82. |
| [3] | Deng J Y. The fundamentals of Ecology of proliferation and management of fishery resources in the Bohai Sea. Progress in Fishery Sciences, 1988, (9): l-10. |
| [4] | Deng J Y, Meng T X, Ren S M, Qiu X Y, Zhu J Y. The composition of fish species and quantity distribution in the Bohai Sea. Progress in Fishery Sciences, 1998, 9(1): 11-89. |
| [5] | Zhu X H, Yang J M, Tang Q S. Study on characteristic of fish community structure in Bohai Sea. Oceanologia Et Limnologia Sinica, 1996, 27(1): 6-13. |
| [6] | Deng J Y, Meng T X, Ren S M. Food web of fishes in the Bohai Sea. Marine Fisheries Research, 1988, (9): 151-171. |
| [7] | Zhu X H, Miao F, Liu D, Xian W W. Spatiotemporal pattern and dominant component of fish community in the Yellow River estuary and its adjacent waters. Studia Marina Sinica, 2001, 43(1): 141-151. |
| [8] | Wang G C, Wang Q. The Comprehensive rehabilitation of fishery resources in Laizhou Bay of Bohai sea. Shandong Fisheries, 2008, 25(10): 43-44. |
| [9] | Li F, Lü Z B, Wei Z H, Wang T T, Xu B Q, Wang Z Q. Seasonal changes in the community structure of the demersal fishery in Laizhou Bay. Journal of Fishery Sciences of China, 2013, 20(1): 137-147. |
| [10] | Tian M C, Sun B L, Yang J M. Analysis of the fish fauna of the Bohai Sea. Studia Marina Sinica, 1993, 34: 157-167. |
| [11] | Deng J Y, Jin X S. Dynamic characteristics of abundance and community structure of fishery species in the overwintering ground of the Bohai Sea. Journal of Natural Resources, 2001, 16(1): 42-46. |
| [12] | Pinkas L, Oliphant M S, Iverson I L K. Food habits of albacore, bluefish tuna, and bonito in California waters. Water Research, 1971, 18(6): 653-594. |
| [13] | Cheng J S. The structure and diversity of demersal fish communities in winter in the East China Sea and the Yellow Sea. Progress in Fishery Sciences, 2000, 21(3): 1-8. |
| [14] | Clarke K R. Comparisons of dominance curves. Journal of Experimental Marine Biology and Ecology, 1990, 138(1): 143-157. |
| [15] | Margalef D R. Information theory in ecology. International Journal of General Systems, 1958, 3(1): 36-71. |
| [16] | Shannon C E, Weaver W. The mathematical theory of Communication. Urbana: University of Illinois Press, 1949, 19(7): 1. |
| [17] | Pielou E C. Ecological Diversity. New York: John Wiley and Sons, 1975. |
| [18] | Bray T R, Curtis J T. An ordination of the upland forest communities of southern Wisconsin. Ecological. Monographs, 1957, 27(4): 325-349. |
| [19] | Ryder R A, S R Kerr, W W Taylor, P A Larkin. Community consequence of fish stock diversity. Canadian Journal of Fisheries and Aquatic Sciences, 1981, 38(12): 1856-1866. |
| [20] | Jin X S, Zhao X Y, Meng T X, Cui Y. Living Resources and Environment in the Yellow Sea and Bohai Sea. Beijing: Science Press, 2005: 262-351. |
| [21] | Yan Y P. Assessment of pollution of heavy metals of economic fish in the coastal waters of China. Marine Environmental, 1993, 12(3/4): 99-103. |
| [22] | Cheng J S, Qiu S Y, Li P J, Cui Y. Ecological environment and biotic community in the coastal waters of the Yellow Sea and Bohai Sea. Qingdao, China Ocean University Press, 2004: 3-190. |
| [23] | Xu Z L, Chen J J. Analysis on migratory routine of Larimichthy polyactis. Journal of Fishery Sciences of China, 2009, 16(6): 931-940. |
| [24] | Zhu X H, Wu H Z, Xu F S, Ye M Z, Zhao Z J. The changes of community diversity of nekton and their correlative factors in the coastal waters of Yellow-Bohai Sea. Acta Oceanologica Sinica, 1994, 16(3): 102-112. |
| [25] | Ding P X, Wang H J, Meng X W, Zhu J R. Evolution and cause analysis of typical coastal zones in China during the last 50 years. Beijing: Science Press, 2013: 264-265. |
| [26] | Luo B Z, Shen H T. The Three Gorges Dam and ecological environment in estuary. Beijing: Scientific Press, 1994: 253-343. |
| [27] | Li J S, Li S F, Ding F Y, Cheng J H. Analysis on annual change of fish diversity in Yangtze estuary off shore water area. Journal of Fishery Sciences of China, 2007, 14(4): 637-643. |
| [28] | Jin X S, Tang Q S. Changes in fish species diversity and dominant species composition in the Yellow Sea. Fisheries Research, 1996, 26(3): 337-352. |
| [29] | Jin X S. The change of community structure of nekton in the waters off southern Shandong Peninsula in spring. Journal of Fisheries of China, 2003, 27(1): 19-24. |
| [30] | Jin X S. Fishery biodiversity and community structure in the Yellow and Bohai Seas. American Fisheries Society Symposium, 2003, 38: 643-650. |
| [31] | Sun Z Z, Qi J Y, Zeng Z C, Zhang Y P, Sun J, Hao Y M. Polychlorinated biphenyls in the water, soil and aquatic animals from the Jiuduansha Wetland of Yangtze River Estuary. Research of Environmental Sciences, 2008, 21(3): 92-97. |
| [32] | Xian W W, Kang B, Liu R Y. Jellyfish Blooms in the Yangtze Estuary. Science, 2005, 307: 41-41. |
| [33] | Tong J A. The main source and distribution characteristics of pollutants in Laizhou Bay. Journal of oceanography of Huanghai and Bohai Seas, 1994, 12(4): 16-20. |
| [34] | Liu Y H, Yang X L, Jin Y, Tang X C, Bai Y Y, Sun S, Tao H M, Gu W L, Wang G H. Distribution and inter-annual variation of nutrients in Laizhou Bay. Progress in Fishery Sciences, 2011, 32(4): 1-5. |
| [35] | Zhang B, Li Z Y, Jin X S. Functional groups of fish assemblages and their major species in the Bohai Sea. Journal of Fisheries of China, 2012, 36(1): 64-72. |
| [36] | Washington H G. Diversity biotic and similarity indices: a review with special relevance to aquatic ecosysterms. Water Research, 1984, 18(6): 653-694. |
| [37] | Jin X S, Deng J Y. Yearly variations of fishery resources and biodiversity in the Laizhou Bay. Progress in Fishery Sciences, 1999, 20(1): 6-12. |
| [38] | Fei H N, He B Q, Chen G M. The regional and seasonal variations of diversity and dominant species of demersal fish communities in continental shelf of northern Nan Hai. Journal of Fisheries of China, 1981, 5(1): 1-20. |
| [39] | Shen J A, Cheng Y H. A study on the deep sea demersal fish communities and their structures in the East China Sea. Journal of Fisheries of China, 1987, 11(4): 293-306. |
| [40] | Wilhm J L. Use of biomass units in shannons formula. Ecology, 1968, 49: 153-156. |
| [41] | Huston M. A general hypothesis of species diversity. The American Naturalist, 1979, 113(1): 81-101. |
| [42] | Jin X S, Shan X J, Guo X W, Li X S. Community structure of fishery biology in the Yangtze River estuary and its adjacent waters. Acta Ecologica Sinica, 2009, 29(9): 4761-4772. |
| [1] | 金显仕, 邓景耀. 莱州湾渔业资源群落结构和生物多样性的变化. 生物多样性, 2000, 8(1): 65-72. |
| [2] | 邓景耀, 金显仕. 莱州湾及黄河口水域渔业生物多样性及其保护研究. 动物学研究, 2000, 21(1): 76-82. |
| [3] | 邓景耀. 渤海渔业资源增殖与管理的生态学基础. 渔业科学进展, 1988, (9): 1-10. |
| [4] | 邓景耀, 孟田湘, 任胜民, 邱显寅, 朱建元. 渤海鱼类种类组成及数量分布. 渔业科学进展, 1998,9(1): 11-89. |
| [5] | 朱鑫华, 杨纪明, 唐启升. 渤海鱼类群落结构特征的研究. 海洋与湖沼, 1996, 27(1): 6-13. |
| [6] | 邓景耀, 孟田湘, 任胜民. 渤海鱼类的食物关系. 渔业科学进展, 1988,(9): 151-171. |
| [7] | 朱鑫华, 缪锋, 刘栋, 线薇薇. 黄河口及邻近海域鱼类群落时空格局与优势种特征研究. 海洋科学集刊, 2001, 43(1): 141-151. |
| [8] | 王广成, 王权. 渤海莱州湾渔业资源综合修复初探. 齐鲁渔业, 2008, 25(10): 43-44. |
| [9] | 李凡, 吕振波, 魏振华, 王田田, 徐炳庆, 王忠全. 2010年莱州湾底层渔业生物群落结构及季节变化. 中国水产科学, 2013, 20(1): 137-147. |
| [10] | 田明诚, 孙宝龄, 杨纪明. 渤海鱼类区系分析.海洋科学集刊, 1993, 34: 157-167. |
| [11] | 邓景耀, 金显仕. 渤海越冬场渔业生物资源量和群落结构的动态特征. 自然资源学报, 2001, 16(1): 42-46. |
| [13] | 程济生. 东、黄海冬季底层鱼类群落结构及其多样性. 渔业科学进展, 2000, 21(3): 1-8. |
| [20] | 金显仕, 赵宪勇, 孟田湘, 崔毅. 黄、渤海生物资源与栖息环境. 北京: 科学出版社, 2005: 262-351. |
| [21] | 闫雨平. 中国近海海域经济鱼类的重盒属污染及其评价. 海洋环境科学, 1993, 12(3/4): 99-103. |
| [22] | 程济生, 邱盛尧, 李培军, 崔毅. 黄渤海近岸水域生态环境与生物群落. 青岛: 中国海洋大学出版社, 2004: 3-190. |
| [23] | 徐兆礼, 陈佳杰. 小黄鱼洄游路线分析. 中国水产科学, 2009, 16(6): 931-940. |
| [24] | 朱鑫华, 吴鹤洲, 徐凤山, 叶懋中, 赵紫晶. 黄渤海沿岸水域游泳动物群落多样性及其相关因素的研究. 海洋学报, 1994, 16(3): 102-112. |
| [25] | 丁平兴, 王厚杰, 孟宪伟, 朱建荣. 近50年我国典型海岸带演变过程与原因分析. 北京: 科学出版社, 2013: 264-265. |
| [26] | 罗秉征, 沈焕庭.三峡工程与河口生态环境. 北京:科学出版社, 1994: 253-343. |
| [27] | 李建生, 李圣法, 丁峰元, 程家骅. 长江口近海鱼类多样性的年际变化.中国水产科学, 2007, 14(4): 637-643. |
| [29] | 金显仕. 山东半岛南部水域春季游泳动物群落结构的变化. 水产学报, 2003, 27(1): 19-24. |
| [31] | 孙振中, 戚隽渊, 曾智超, 张玉平, 孙骥, 郝永梅. 长江口九段沙水域环境及生物体内多氯联苯分布. 环境科学研究, 2008, 21(3): 92-97. |
| [33] | 童钧安. 莱州湾主要污染物来源及分布特征. 黄渤海海洋, 1994, 12(4): 16-20. |
| [34] | 刘义豪, 杨秀兰, 靳洋, 汤宪春, 白艳艳, 孙珊, 陶慧敏, 谷伟力, 王国华. 莱州湾海域营养盐现状及年际变化规律. 渔业科学进展, 2011, 32(4): 1-5. |
| [35] | 张波, 李忠义, 金显仕. 渤海鱼类群落功能群及其主要种类. 水产学报, 2012, 36(1): 64-72. |
| [37] | 金显仕, 邓景耀. 莱州湾春季渔业资源及生物多样性的年间变化. 渔业科学进展, 1999, 20(1): 6-12. |
| [38] | 费鸿年, 何宝全, 陈国铭. 南海北部大陆架底栖鱼群聚的多样度以及优势种区域和季节变化. 水产学报, 1981, 5(1): 1-20. |
| [39] | 沈金鏊, 程炎宏. 东海深海底层鱼类群落及其结构的研究. 水产学报, 1987, 11(4): 293-306. |
| [42] | 金显仕, 单秀娟, 郭学武, 李显森. 长江口及其邻近海域渔业生物的群落结构特征. 生态学报, 2009, 29(9): 4761-4772. |