 2014, Vol. 34
2014, Vol. 34文章信息
- 谭秋锦, 宋同清, 彭晚霞, 曾馥平, 杨钙仁, 杜虎, 鹿士杨, 范夫静
- TAN Qiujin, SONG Tongqing, PENG Wanxia, ZENG Fuping, YANG Gairen, DU Hu, LU Shiyang, FAN Fujing
- 西南峡谷型喀斯特不同生态系统的碳格局
- Carbon pattern of different ecosystems in canyon karst region in southwestern of China
- 生态学报, 2014, 34(19): 5579-5588
- Acta Ecologica Sinica, 2014, 34(19): 5579-5588
- http://dx.doi.org/10.5846/stxb201301150101
-
文章历史
- 收稿日期:2013-1-15
- 网络出版日期:2014-3-7






2. 广西大学林学院, 南宁 530004;
3. 中国科学院环江喀斯特生态系统观测研究站, 环江 547100;
4. 江西农业大学, 南昌 330045
2. Forestry College of Guangxi University, Nanning 530004, China;
3. Huanjiang Observation and Research Station of Karst Ecosystem, Chinese Academy of Sciences, Huanjiang, Guangxi Zhuang Autonomous Region, Huanjiang 547100, China;
4. Jiangxi Agricultural University, Nanchang, Jiangxi 330045, China
随着大气CO2浓度升高,全球气候环境问题日益严峻,且今后很长一段时期内不可能实现零排放,大气CO2浓度增高不可能逆转[1]。其中农田、草地、森林3类主要陆地生态系统中的碳库贮量被认为是一种非常有效的CO2减排措施,农田土壤碳库的稳定与增加对于保证全球粮食安全与缓解气候变化趋势具有双重积极意义[2]。许多研究表明,发达国家实行的保护性耕作、秸秆还田、施用有机肥和化肥等农业管理措施,使得农业土壤碳库在近期呈现出稳定和增长的趋势[3]。草地生态系统覆盖了几乎20%的陆地面积,其碳贮量占全球陆地生态系统1/3[4],Gurney和Neff评估了美国、加拿大和俄罗斯草地改良的固碳潜力[5],IPCC估算全球草地2010年的固碳潜力为0.24Pg[6]。森林维持的植被碳库约占全球植被碳库的86%[7]。维持的土壤碳库约占全球土壤碳库的73%[8]。同时,森林生态系统具有较高的生产力,每年固定的有机碳约占整个陆地生态系统的 2/3[9, 10],其中地上部分占80%,地下部分占40%[11]。 王斌和杨校生[12]对西双版纳热带季节雨林、鼎湖山亚热带常绿阔叶林、哀牢山中山湿性常绿阔叶林和长白山阔叶红松林4 种地带性森林类型的碳含量和碳密度及其空间分布格局进行了比较,Chen等[13]研究了中国南方马尾松林、杉木林、硬阔林和软阔林乔木层地上生物量及其碳汇能力,指出阔叶林是我国热带和亚热带区域碳汇林的最佳选择。
喀斯特生态系统是以碳酸盐为主要的物质基础,在大气-水-岩-生物相互作用下所形成的一种具有特殊物质、能量、结构和功能的生态系统,具有景观异质性强、二元结构明显、环境容量小、植被不连续、土层浅薄、蓄水性差、土地承载力小、抗干扰能力弱等特点[14]。近年来,在人为活动的干扰加剧情况下,产生了不同程度的退化,许多地方石漠化现象严重,且仍呈扩张的态势。目前对喀斯特植被群落的退化原因、退化过程及与环境的关系研究较多[15, 16, 17, 18, 19],但有关喀斯特地区碳储量特别是同一区域不同生态系统碳格局的比较研究报道甚少[20],本文以贵州省晴隆县孟寨小流域峡谷型喀斯特的6个生态系统为研究对象,根据样地调查数据估算各生态系统碳格局并进行了比较,旨在揭示不同生态系统总体生物量碳库和土壤碳库的变化规律,为制定该区域的植被恢复、生态重建和固碳措施提供科学参考。
1 研究区概况与研究方法 1.1 研究区概况研究区为贵州省晴隆县孟寨河小流域,位于25°33′—26°11′N、105°01′—105°25′E之间,最高海拔2025 m,属北亚热带湿润季风气候区,年平均气温14.0—15.9℃,年极端高温为33.4℃,最低气温为-6.2℃,日照时数1 453 h,无霜期280 d。年降水量1500—1650 mm,集中在6—9月份,年蒸发量1 800 mm,空气相对湿度在50%以下。地形起伏大,具有山高、谷深、坡陡的特点,属二迭纪岩层风化而成的石灰土。该小流域与(高原型)普定后寨、(岩溶槽谷型)印江朗溪、 (峰丛洼地型)荔波板寨并称四大地貌类型区的4个小流域。
在流域内选取立地条件基本一致、具有代表性的水田、旱地、草地、灌丛、人工林和次生林6种生态系统,其中水田种植以水稻(Oryza sativa)、小麦(Triticum aestivum)为主,旱地种植玉米(Zea mays)、油菜(Brassica napus)等;人工草地的主要植物组成有白花刺(Sophora davidii)、宽叶雀稗(Paspalum wettsteinii Hackel)、多年生黑麦草(Lolium perenne)、鸭茅(Dactylis glomerata)、白三叶(Trifolium repens)、紫花苜蓿(Medicago sativa)等,草地上饲养的家畜主要是山羊,品种包括杂交波尔山羊(Transgressus Boer Capra)、本地黑山羊(Local niger hircum)、南江黄羊(Nanjiang Yellow)和杜泊羊(Dorper oves),以放牧利用为主;灌丛主要有车桑子(Dodonaea viscosa (Linn.) Jacq. Enum)、高羊茅(Festucaarundinacea)等;人工林主要是人为种植的楸树(Catalpa bungei C.A.Mey.),林龄在15—20 a之间,群落结构简单,林下植被发育较差且分布不均,盖度仅6%左右;灌木层主要有火棘(Pyracantha fortuneana)、杜茎山(Elaeocarpus syluestris)等;次生林林龄20—40 a,乔木层以白栎(Quercus fabri Hance)、椿树(Toona sinensis)和翅荚香槐(Cladrastis platycarpa)为优势树种,灌木层植物主要包括朴树(Celtis sinensis Pers)、构树(Broussonetia papyrifera)、香叶树(Lindera communis Hemsl)和阴香(Cinnamomum burmannii)等。
1.2 研究方法 1.2.1 样方设置与调查对研究区域进行实地踏查基础上,选择具有代表性、立地条件基本一致的水田(ST)、旱地(HD)、草地(CD)、灌丛(GC)、人工林(RGL)、次生林(CSL)6个生态系统,各设置3个样地(样地特征见表 1),样地面积为20 m×20 m。乔木调查主要包括种名、DBH、树高、冠幅和存活状态,并进行每木检尺,起测胸径(DBH)为5 cm;灌木层按“品”字型选择3个2 m×2 m的样框,调查种名、株丛数、平均高度、平均基径和盖度;草本层在灌木层样框内各选择1个1 m×1 m的样框,调查其种名、株丛数、平均高度,并记录覆盖度,同时用GPS定位。
| 生态系统 Ecosystems | 坡度 Slope / (°) | 土壤类型 Soil type | 土壤深度 Soil depth /cm | 平均胸径 Mean DBH /cm | 平均年龄 Mean age /a | 平均高度 Mean height /m | 郁闭度 Coverage |
| ST:水田Paddy field;HD:旱地Dry land;CD:草地GC: shrubber;GC:灌草GC: shrubbery;RGL:人工林artificial forest;CSL:次生林;Secondary forest | |||||||
| ST | 0 | 棕色石灰土 | 45.3 | ||||
| HD | 8 | 棕色石灰土 | 49.0 | ||||
| CD | 2 | 棕色石灰土 | 53.9 | 2 | 0.2 | 0.95 | |
| GC | 17 | 棕色石灰土 | 18.9 | 0.6 | 3 | 1.3 | 0.75 |
| RGL | 22 | 棕色石灰土 | 19.2 | 10.6 | 12 | 7.0 | 0.5 |
| CSL | 25 | 棕色石灰土 | 30.0 | 9.72 | 20 | 7.8 | 0.7 |
乔木层:在每木调查的基础上,以2 cm为径阶,根据中央径阶多、两端逐次少的原则,在人工林样地内选择楸树15株、次生林样地内选择白栎10株、椿树3株、翅荚香槐2株作为样木,进行生物量的测定。测定其基径、胸径、树高和冠幅,并按2 m区分段锯断称量,在树干基部、胸径、中部及顶端分别锯取一个圆盘测定含水量;并分别称取枝条、叶片和花果的鲜质量;根的生物量采用全挖法,并分小根(<2 cm)、中根(2—5 cm)、大根(>5 cm)3组分别称重;然后对各部分器官分别取样500 g带回实验室,将样品在105 ℃烘箱内杀青2 h,调至75 ℃下烘干至恒重,求出各器官干鲜质量之比,由此换算出样木各器官的干质量及总干质量。利用样木各器官(干、枝、叶、根)的干重和总干重建立如表 2所示的各器官及总生物量与样木胸径(D)的幂回归方程(W=aDb)。根据样地每木调查的结果及相应的回归方程计算乔木层各个体的生物量,并由此获得乔木层的总生物量。
| 类型Types | 拟合方程 Fitting equation | 相关系数R2 | 残差平方和Sum of squared residuals |
| ** P<0.01 | |||
| 人工林Artificial forest | Y=0.261D2.475 | 0.967* * | 0.291 |
| 次生林Secondary forest | Y=0.179D2.335 | 0.945* * | 1.820 |
灌木层、草本层和地上凋落物:在灌木层样框内按全收获法收获样框内所有灌木的枝、叶、花果、根,测定各器官生物量,并以3个样框的平均值来推算样地中灌木层的总生物量。按同样方法在1 m×1 m草本小样框内获取草本层地上与地下、凋落物生物量及总生物量(调查时农田和旱地生态系统地上部分已刈割)。
1.2.3 土样采集首先将每个生态系统样地划分为16个5 m×5 m的小样方,各获得25个网点,用土钻测定每个网格点的土壤厚度。在每个样地中挖取典型剖面,直至出现岩石层为止,用环刀分0—10 cm(Ⅰ层)、10—20 cm(Ⅱ层)、20—30 cm(Ⅲ层)、30—50 cm(Ⅳ层)、50—100 cm(Ⅴ层)5层测容重。在样地的4个角和中间按土壤机械分5层取样,5个点同层次的土样组成一个混合样。样品取完后置于阴凉处自然风干,用四分法取土样过筛,供分析测定用,同时测定石砾含量。
1.2.4 样品分析植物碳和土壤碳均采用重铬酸钾-浓硫酸氧化外加热法测定全碳含量[21]。
1.2.5 碳贮量计算林下植被(灌木层、草本层)和凋落物层的碳贮量以其生物量现存量乘以相应的碳含量求得。土壤层碳贮量的计算采用以下公式:
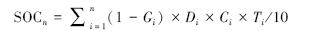
式中,SOCn为分n层调查的土壤单位面积碳贮量(Mg C/hm2),Gi为第i层直径≥2 mm的石砾含量(%),Di为第i层土壤容重(g/cm3),Ci为第i层土壤有机碳含量(g/kg),Ti为第i土层的厚度(cm)。
1.2.6 数据处理采用单因素方差分析和多因素方差分析分析6种不同生态系统碳含量、碳贮量的空间变异情况,并采用最小显著差法进行多重比较。所应用的软件为Excel-2007、SPSS 16.0。
2 结果与分析 2.1 植物碳贮量采样时旱地和水田地上部分已刈割,但是刘阳等[22]研究传统耕作和保护性耕作2种方式对玉米农田生态系统的各碳库的影响,知道地上部分为0.39×103—1.43×103 kg C/hm2,地下部分为0.11×103—0.36×103 kg C/hm2,而李庆云[23]研究间作样地不同时间小麦的碳贮量为2.21×103—2.81×103 kg C/hm2。由表 3知,如以上面的数据作比较,水田和旱地较其它4种生态系统的碳贮量少,而现两者已刈割,只对草地、灌丛、人工林和次生林4种生态系统的碳贮量进行现状比较和分析。植被总碳贮量为RGL>CSL>GC>CD,差异极显著,RGL ∶ CSL ∶ GC ∶ CD为1 ∶ 0.96 ∶ 0.45 ∶ 0.30,均以地上部分为主,地上部分分别是根系的3.51、8.01、1.61、1.49倍, 地上部分的大小顺序为CSL>RGL>GC>CD,根系则为RGL>GC>CD>CSL,灌丛、草地、次生林差异不显著,均极显著低于人工林。同一生态中不同层次的碳储量对植被总碳储量的贡献不同,草地只有草本层,碳储量为36.05×103 kg C/hm2;灌丛没有乔木层,灌木层碳储量的贡献率达87.64%,是草本层的7.10倍;人工林和次生林两类森林生态系统的乔木层碳储量占绝对优势,分别是总碳储量的93.36%和88.05%,其他各层所占比例很小,人工林的草本层>灌木层,次生林反之。同一层次不同生态系统的碳储量不同,乔木层人工林极显著高于次生林;灌木层为GC>CSL>RGL,差异极显著;草本层为草地极显著高于灌丛、人工林、次生林,后三者差异不显著。
| 生态系统 Ecosystems | 乔木层 Tree layer/ (103kg C/hm2) | 灌木层 Shrub layer/ (103kg C/hm2) | 草本层 Herb layer/ (103kg C/hm2) | 合计Total(103kg C/hm2) | ||||||||
| 地上部分 Aboveground | 根 Root | 小计 Total | 地上部分 Aboveground | 根 Root | 小计 Total | 地上部分 Aboveground | 根 Root | 小计 Total | 地上部分 Aboveground | 根 Root | 小计 Total | |
| 同列不同大写字母表示差异极显著(P<0.01),不同小写字母表示差异显著(P<0.05) | ||||||||||||
| CD | 21.59Aa | 14.46Aa | 36.05Aa | 21.59Dd | 14.46Bb | 36.05Dd | ||||||
| GC | 28.93Aa | 18.52Aa | 47.45Aa | 4.44Bb | 2.25Bb | 6.69Bb | 33.37Cc | 20.77Bb | 54.14Cc | |||
| RGL | 88.58Bb | 24.88Aa | 113.46Aa | 1.73Cc | 0.42Cc | 2.15Cc | 4.25Bb | 1.67Bb | 5.92b | 94.56Aa | 26.97Aa | 121.53Aa |
| CSL | 94.99Aa | 7.82Bb | 102.81Bb | 5.64Bb | 3.61Bb | 9.22Bb | 3.17Bb | 1.53Bb | 4.70b | 103.80Bb | 12.96Bb | 116.76Bb |
6种生态系统中,次生林地被物的碳贮量最大,极显著高于灌丛、人工林和草地(P﹤0.01),灌丛和人工林没有显著差异,但都极显著高于草地(表 5),调查时水田和旱地没有地被物。尽管各个生态系统中地被物碳贮量的绝对数量相差较大,但对整个生态系统碳贮量的贡献不大,均低于5%。
2.3 土壤碳峡谷型喀斯特6种典型生态系统土壤碳含量均随土壤深度的增加极显著降低(表 4),其中CSL和CD的下降速度相对平缓,RGL和GC下降的速度最快。从ST到CSL,第Ⅱ层的碳含量依次为第Ⅰ层的72.33%、81.78%、99.32%、55.93%、87.43%和93.10%,第Ⅲ层为第Ⅰ层的50.78%、60.46%、78.85%、34.37%、62.93%和79.37%,最后一层为第Ⅰ层的36.30%、35.43%、50.20%、34.37%、20.31%、79.37%。不同生态系统同一层次的土壤碳含量也有明显差异。第Ⅰ层,CSL和RGL的土壤碳含量差异不显著,但均显著高于GC(P﹤0.05),极显著高于ST、HD、CD(P﹤0.01),GC和ST不显著,均显著高于HD和CD(P﹤0.05),HD和CD之间无显著差异。第Ⅱ层,CSL和RGL的碳含量差异不显著,极显著高于ST、HD、CD和GC(P﹤0.01),后4者间的差异不显著。第Ⅲ层,CSL和RGL差异不显著,但极显著高于CD、ST、HD和GC(P﹤0.01),CD显著高于ST、HD和GC(P﹤0.05),ST显著高于HD和GC(P﹤0.05),HD和GC无显著差异。第Ⅳ层,CT、HD显著高于ST、RGL(P﹤0.05)。第Ⅴ层,ST、HD和CD之间无显著差异。土壤不同层次的碳贮量变化趋势相同(表 4)。
| 生态系统 Ecosystems | 土壤层次 Soil layer | 碳含量 SOC content/ (g C/kg) | 碳贮量 SOC Storage/ (103kg C/hm2) | 土壤平均碳含量 Average SOC content/(g C/kg) | 土壤碳贮量 SOC Storage/ (103kg C/hm2) |
| ST | Ⅰ | 29.42Aa | 35.54Aa | 17.56c | 112.26Bb |
| Ⅱ | 21.28Bb | 29.55Bb | |||
| Ⅲ | 14.94Cc | 22.39Cc | |||
| Ⅳ | 11.50Dd | 24.39Cc | |||
| Ⅴ | 10.68Ee | 0.35Dd | |||
| HD | Ⅰ | 23.98Aa | 30.17Aa | 16.70c | 115.31Bb |
| Ⅱ | 19.61Bb | 25.54Aa | |||
| Ⅲ | 14.50Cc | 24.06Aa | |||
| Ⅳ | 15.00Dd | 29.83Aa | |||
| Ⅴ | 10.40Ee | 5.70Bb | |||
| CD | Ⅰ | 25.34Aa | 36.97a | 19.81c | 153.12Aa |
| Ⅱ | 25.17Bb | 34.85a | |||
| Ⅲ | 19.98Cc | 28.22b | |||
| Ⅳ | 15.85Dd | 40.89Aa | |||
| Ⅴ | 12.72Ee | 17.15c | |||
| GC | Ⅰ | 33.90Aa | 36.23Aa | 21.50c | 52.69Cc |
| Ⅱ | 18.96Bb | 16.23Bb | |||
| Ⅲ | 11.65Cc | 0.23Cc | |||
| RGL | Ⅰ | 43.43Aa | 46.85Aa | 29.39Bb | 65.04Cc |
| Ⅱ | 37.97Bb | 17.10Bb | |||
| Ⅲ | 27.33Cc | 1.01Cc | |||
| Ⅳ | 8.82Dd | 0.08Cc | |||
| CSL | Ⅰ | 48.57Aa | 52.46Aa | 44.11Aa | 126.11Bb |
| Ⅱ | 45.22Bb | 50.30Aa | |||
| Ⅲ | 38.55Cc | 23.35Bb |
6 种生态系统土壤碳的平均含量可分为3组,CSL>RGL>GC、CD、ST、HD,组间差异极显著,组内差异不显著(表 4),其中CSL分别是RGL、GC、CD、ST、HD的1.50、2.05、2.23、2.51、2.64倍,RGL分别是GC、CD、ST、HD的1.37、1.48、1.67、1.76倍。同理,6种生态系统土壤碳贮量也可分为3组,CD>CSL、ST、HD>RGL、GC,组间差异极显著,组内差异不显著(表 4),主要受土壤深度的影响,其中CD土壤碳贮量分别是CSL、ST、HD、RGL、GC的1.21、1.36、1.33、2.35、2.91倍。
2.4 不同生态系统的碳贮存及其分布格局6种生态系统的总碳贮量大小顺序为:CSL>RGL>CD>HD>GC>ST,CSL的碳贮量最高,极显著高于RGL,RGL极显著高于CD,CD极显著高于GC、HD、ST,GC、HD、ST的差异不显著(表 5)。同一生态系统碳贮量的分布格局不同,CSL碳贮量的分布格局为土壤﹥植物﹥地被物,RGL碳贮量的分布格局则为植物﹥土壤﹥地被物,都是地上部分>地下部分;GC的植物和土壤碳储量差异很小,基本平衡,但均明显高于地被物、地下部分碳贮量高于地上部分;CD的碳贮量贡献率为土壤>植物>地被物、地下部分>地上部分;HD和ST调查时地上部分已刈割,无植物和地被物碳储量,其土壤碳贮量分别为115.31×103 kgC/hm2和112.26 ×103 kgC/hm2。
| 类型 Types | 植物 Plant | 地被物层 Ground cover | 土壤 Soil | 地上部分 Aboveground | 地下部分 Underground | 合计 Total | |||||
| 碳贮量 Carbon storage/ (103kg C/hm2) | % | 碳贮量 Carbon storage/ (103kg C/hm2) | % | 碳贮量 Carbon storage/ (103kg C/hm2) | % | 碳贮量 Carbon storage/ (103kg C/hm2) | % | 碳贮量 Carbon storage/ (103kg C/hm2) | % | ||
| 地上部分碳贮量包括地被物碳与植物地上部分所含的碳,地下部分碳贮量则包括土壤碳和根系碳; 碳贮量单位:103kg C/hm2 | |||||||||||
| ST | 0.00 | 0.00 | 0.00 | 0.00 | 112.26Bb | 100.00 | 0.00 | 0.00 | 112.26Dd | 100.00 | 112.26Dd |
| HD | 0.00 | 0.00 | 0.00 | 0.00 | 115.31b | 100.00 | 0.00 | 0.00 | 115.31Dd | 100.00 | 115.31Dd |
| CD | 36.05Dd | 19.01 | 0.47Cc | 0.25 | 153.12Bb | 80.74 | 22.06Dd | 11.63 | 167.58Cc | 88.37 | 189.64Cc |
| GC | 54.14Cc | 48.06 | 5.82Bb | 5.17 | 52.69Cc | 46.77 | 39.19Cc | 34.79 | 73.46Dd | 65.21 | 112.65Dd |
| RGL | 121.53Bb | 63.79 | 3.96Bb | 2.07 | 65.04Bb | 33.14 | 98.52Bb | 51.71 | 92.01Bb | 48.29 | 190.53Bb |
| CSL | 116.76Aa | 44.70 | 18.34Aa | 7.02 | 126.11Aa | 48.28 | 122.14Aa | 46.76 | 139.07Aa | 53.24 | 261.21Aa |
西南喀斯特植物在区划上属泛北极植物区,中国-日本森林植物区,滇黔桂地区范畴[24, 25],其顶极群落为亚热带喀斯特常绿落叶阔叶混交林,生境异质性极高、区系组分复杂,生物种类极为丰富,生态系统稳定。但受地球内动力、强烈的地质运动、高温多雨且分布不均、碳酸盐岩溶蚀性强、水文二维结构明显以及其适生植物具有嗜钙性、耐旱性和石生性等限制特点的综合影响,生态系统的稳定性和抗干扰能力差[14],强烈的人为干扰导致农田、旱地、草丛、灌丛、人工林、次生林、原生林等各种偏途、不同演替阶段生态系统和群落共存现象。峡谷型喀斯特区域人口密度大,干扰更严重,顶极群落破坏殆尽。植被碳储量为36.05×103—121.53×103kg C/hm2,其大小顺序为RGL>CSL>GC>CD,差异极显著,同区域喀斯特峰丛洼地石灰土上的相同生态系统相似(5.02×103—136.17×103kg C/hm2),如次生林的植物碳贮量高达116.76×103kg C/hm2,远高于峰丛洼地石灰土上的次生林和红壤上的白桦林、马尾松落叶阔叶混交林[26, 27, 28](表 6)。不同生态系统植被各层次的碳储量贡献不同,次生林和人工林两类乔木层的碳贮量占绝对优势,分别为总碳贮量的93.36%和88.05%,灌丛的灌木层碳贮量贡献高达87.64%,草地的草本层碳贮量贡献率为100%。同一层次不同生态系统碳贮量也不同,乔木层人工林极显著高于次生林;灌木层为GC>CSL>RGL,差异极显著;草本层为草地极显著高于灌丛、人工林、次生林。
| 森林类型 Forest types | 地点 Sites | 土层厚度 Soil thickness/ (103kg C/hm2) | 土壤碳Soil carbon/ (103kg C/hm2) | 植被碳贮量 Vegetation carbon storage/ (103kg C/hm2) | 文献 References | |
| 含量 Content | 贮量 Storage | |||||
| 峡谷型次生林 Secondary forest in Canyon type | 贵州省晴隆县 | 30 | 44.11 | 126.11 | 229.61 | |
| 峰丛洼地次生林 Secondary forest in Cluster-peak- depression Region | 广西区环江县 | 60 | 33.29 | 121.43 | 60.19 | [26] |
| 白桦林 Bemla platyphlla plantation | 四川省平武县 | 60 | 22.64 | 165.89 | 73.74 | [27] |
| 中亚热带常绿阔叶林 Evergreen broad-leaved forests in mid-subtropical | 湖南省会同县 | 60 | 20.08 | 65.73 | 110.98 | [28] |
西南峡谷型喀斯特受亚热带湿润季风气候影响较大,优越的温湿条件既有利于生物的繁衍和生长,增加生物“自肥”作用,同时加速了岩石的溶蚀、风化和土壤的形成与发育进程[29]。土壤有机碳含量为16.70×103—44.11×103kg C/hm2,远高于同纬度地区红壤[30],以次生林为例,其含量高达44.11×103kg C/hm2,远高于同纬度地区红壤上的白桦林和马尾松落叶阔叶混交林,同时也高于同纬度地区峰丛洼地石灰土上的次生林(表 6)。土壤碳贮量略低于同纬度地区红壤上的白桦林,但远高于同纬度地区峰丛洼地次生林和马尾松落叶阔叶混交林。不同生态系统土壤碳含量和碳储量大小不同,碳含量的大小顺序为CSL>RGL >GC>CD>ST>HD,碳储量的大小顺序为CD>CSL>HD>ST>RGL>GC,且土壤碳含量与碳贮量均随深度的增加而显著的降低,且降低程度与土壤深度(层次)有较好的线性关系,土壤碳含量R2的范围在0.891与0.960之间;土壤碳贮量R2的范围在0.823与0.955之间(Y=aX+b,式中Y为土壤碳含量或碳贮量,X为土壤层次,a和b为拟合参数;表 7),其原因一是表层凋落物多,有机质分解量大,二是在野外采集样品时发现表层的细根要明显多于下层,凋落物的增加、根系的垂直分布及其穿插作用直接影响到土壤剖面各层次的有机碳[31]。可见,小尺度上土壤碳贮量主要取决于土层厚度和植物群落,而在更大的尺度上,则还与土壤、气候、土壤理化性质、人为干扰等因子有关。
| 生态系统 Ecosystems | 土壤碳含量 SOC content/(g C/kg) | R2 | 土壤碳贮量 Carbon storage/(103kg C/hm2) | R2 |
| ST | Y=31.742-4.726X | 0.910 | Y=39.556-5.164X | 0.947 |
| HD | Y=26.225-3.175X | 0.927 | Y=32.652-3.116X | 0.955 |
| CD | Y=30.180-3.456X | 0.953 | Y=42.912-5.000X | 0.903 |
| GC | Y=36.198-6.914X | 0.891 | Y=36.806-6.652X | 0.823 |
| RGL | Y=56.569-10.729X | 0.960 | Y=60.433-10.881X | 0.937 |
| CSL | Y=53.857-5.043X | 0.943 | Y=59.243-5.263X | 0.869 |
峡谷型喀斯特6种生态系统的总碳贮量为112.26—261.21×103 kg C/hm2,其大小顺序为CSL>RGL>CD>HD>GC>ST,其中群落发育程度较高的次生林的总碳贮量高达261.21×103 kg C/hm2,高于同区域喀斯特峰丛洼地上次生林和红壤上的白桦林、马尾松落叶阔叶混交林,但远低于红壤上的岷江冷杉林(618.86×103 kg C/hm2)和紫果云杉林(729.92×103 kg C/hm2)[13, 14, 15],不同生态系统碳储量的分布格局不同,地被物的贡献最低,仅为0.25×103—4.90×103 kg C/hm2,次生林和人工林的碳贮量为植物大于土壤,地上大于地下,灌丛的植物和土壤碳贮量相似,地下高于地上,草地、旱地、水田均为土壤大于植物,地下大于地上。
本研究虽然计算了地被物层的碳贮量,涉及到不同植物层次和地下土壤根系,但没有涉及到土壤微生物和土壤动物碳贮量、C周转、群落结构的变化等,即本文所研究的是“某一个时间点”的生态系统碳贮量,其大小顺序为CSL>RGL>CD>GC>HD>ST。其中,次生林的土壤碳含量最高,为44.11 g C/kg;而人工林的植物碳贮量最高,为121.53×103 kg C/hm2;并且各生态系统土壤碳贮量与土壤碳含量均以土壤深度之间有良好的线性关系。实际上,生态系统的碳贮量将随时间而发生相应的变化。不同生态系统碳贮量通常存在较大差异,受研究条件限制,各生态系统类型的立地因子(如海拔、坡度、坡位)和主要林分因子(如林分年龄)存在较大差异,使结果的可比性较低。此外,样地和标准木数量限制也可能会导致碳贮存评估的精确度降低。今后研究中应考虑土壤微生物、群落结构动态所导致的碳贮量变化,尽量获取大量有代表性的生态系统碳贮量实测数,减少区域尺度碳平衡研究中的不确定性,从而为区域碳贮量的精确评估提供充足的基础数据。
本研究初步发现峡谷型喀斯特生态系统碳贮存潜力很大,如改造水田和旱地的单一模式,实行间作耕种模式确保一年四季都有适合其的植物生长;适当对草地和灌丛施加有机肥,提高其植物生长速度,增加碳贮量。总之合适的树种及其搭配比例、退耕还草造林模式和生态系统管理对策等措施,是促进该区域植被恢复和增加碳贮存的关键,同时对减缓全球大气CO2 浓度升高也有着重要意义。
| [1] | Ding Z L, Fu B J, Han X G, Ge Q S. Brief introduction to a cluster of projects of "Research in key issues of international negotiation with regard to coping with climate change" by CAS. Bulletin of the Chinese Academy of Sciences, 2009, 24(1): 7-17. |
| [2] | Li C S, Xiao X M, Frolking S, Moore Ⅲ B, Salas W, Qiu J J, Zhang Y, Zhuang Y H, Wang X K, Dai Z H, Liu J Y, Qin X G, Liu B H, Sass R. Greenhouse gas emissions from croplands of China. Quaternary Sciences, 2003, 23(5): 493-503. |
| [3] | Smith P. Carbon sequestration in croplands: the potential in Europe and the global context. European Journal of Agronomy, 2004, 20(3): 229-236. |
| [4] | Chen Z Z. Typical Steppe Ecosystem in China. Beijing: Science Press, 2000. |
| [5] | Gurney K, Neff J. Carbon Sequestration Potential in Canada//Russia and the United States under Article 3. 4 of the Kyoto Protocol, World Wildlife Fund. 2000. |
| [6] | Metz B, Davidson O, Swart R, Pan J H. Climate Change 2001, Mitigation 2001: Migration: Contribution of Working Group III to the Third Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press, 2001. |
| [7] | Woodwell G M, Whittaker R H, Reiners W A, Likens G E, Delwiche C C, Botkin D B. The biota and the world carbon budget. Science, 1978, 199(4325): 141-146. |
| [8] | Post W M, Emanuel W R, Zinke P J, Stangenberger A G. Soil carbon pools and world life zones. Nature, 1982, 298(5870): 156-159. |
| [9] | Kramer P J. Carbon dioxide concentration, photosynthesis, and dry matter production. Bio Science, 1981, 31(1): 29-33. |
| [10] | Waring R H, Schlesinger W H. Forest Ecosystems: Concepts and Management. London: Academic Press, 1985. |
| [11] | Malhi Y, Baldocchi D D, Jarvis P G. The carbon balance of tropical, temperate and boreal forests. Plant Cell and Environment, 1999, 22(6): 715-740. |
| [12] | Wang B, Yang X S. Comparison of carbon content and carbon density of four typical zonal forest ecosystems. Journal of Hunan Agricultural University: Science and Technology, 2010, 36(4): 465-469. |
| [13] | Chen Q Q, Xu W Q, Li S G, Fu S L, Yan J H. Aboveground biomass and corresponding carbon sequestration ability of four major forest types in south China. Chinese Science Bulletin, 2013, 58(13): 1551-1557. |
| [14] | Peng W X, Wang K L, Song T Q, Zeng F P, Wang J R. Controlling and restoration models of complex degradation of vulnerable Karst ecosystem. Acta Ecologica Sinica, 2008, 28(2): 0811-0820. |
| [15] | Peng W X, Song T Q, Zeng F P, Wang K L, Fu W, Liu L, Du H, Lu S Y, Yin Q C. The coupling relationships between vegetation, soil, and topography factors in karst mixed evergreen and deciduous broadleaf forest. Acta Ecologica Sinica, 2010, 30(13): 3472-3481 |
| [16] | Song T Q, Peng W X, Zeng F P, Wang K L, Qin W G, Tan W N, Liu L, Du H, Lu S Y. Spatial pattern of forest communities and environmental interpretation in Mulun National Nature Reserve, karst cluster-peak depression region. Chinese Journal of Plant Ecology, 2010, 34(3): 298-308. |
| [17] | Song T Q, Peng W X, Zeng F P, Wang K L, Cao H L, Li X K, Qin W G, Liu L. Community composition and biodiversity characteristics of forests in Karst cluster-peak-depression region. Biodiversity Science, 2010, 18(4): 355-364. |
| [18] | Zeng F P, Peng W X, Song T Q, Wang K L, Wu H Y, Song X J, Zeng Z X. Changes in vegetation after 22 years' natural restoration in the karst disturbed area in Northwest Guangxi. Acta Ecologica Sinica, 2007, 27(12): 5110-5119. |
| [19] | Liang S C. Study on dynamics of pubescent hornbeam population in karst mountain of Guiyang. Acta Ecologica Sinica, 1992, 12(1): 53-60. |
| [20] | Song M, Peng W X, Zou D S, Zeng F P, Du H, Lu S Y. The causes of spatial variability of surface soil organic matter in different forests in depressions between karst hills. Acta Ecologica Sinica, 2012, 32(19): 6259-6269. |
| [21] | Bao S D. Soil Agrochemical Analysis. Beijing: Agriculture Publishing House, 1988: 33-36. |
| [22] | Liu Y, Wang J, Liu J, Chi Y C, Zhang X W, Zhao P L. Effect of conservation tillage on carbon storage in corn fields. Inner Mongolia Forestry Science and Technology, 2012, 38(1): 14-17. |
| [23] | Liu Q Y. The Research on Carbon Storage of Populus-Crop Intercropping System in the Huanghuaihai Plain [D]. He'nan: He'nan Agricultural University, 2008. |
| [24] | Wu Z Y. Notes on the distribution patterns of world families of seed plants. Acta Botanica Yunnanica, 2003, 25(5): 535-538. |
| [25] | Wu Z Y, Zhou Z K, Li D Z, Peng H, Sun H. The areal-types of the world families of seed plants. Acta Botanica Yunnanica, 2003, 25(3): 245-257. |
| [26] | He N. The Allocation of Carbon Storage and Its Formation Mechanism of Forest Ecosystems in Depressions between Karst Hills [D]. Hunan: Institute of Subtropical Agriculture, the Chinese Academy of Science, 2012. |
| [27] | Gong C, Wang S L, Zeng Z Q, Deng S J, Chen J P, Long K S. Carbon storage and its distribution pattern of evergreen broad-leaved forests at different succession stages in mid-subtropical China. Chinese Journal of Ecology, 2011, 30(9): 1935-1941. |
| [28] | Xian J R, Zhang Y B, Wang K Y, Hu T X, Yang H. Carbon Stock and its allocation in five forest ecosystems in the subalpine coniferous forest zone of western Sichuan Province, Southwest China. Chinese Journal of Plant Ecology, 2009, 33(2): 283-290. |
| [29] | Han M R, Song T Q, Peng W X, Huang G Q, Du H, Lu S Y, Shi W W. Compositional characteristics and roles of soil mineral substances in depressions between hills in karst region. Chinese Journal of Applied Ecology, 2012, 23(3): 685-693. |
| [30] | Song T Q, Wang K L, Peng W X, Wang J R, Xiao R L, Zeng F P, Tang Y. Ecological effects of intercropping white clover on tea plantation in a subtropical hilly region. Acta Ecologica Sinica, 2006, 26(11): 3647-3655. |
| [31] | Liu X L, Ma Q Y, Yang D S, Shi Z M, Su Y M, Zhou S Q, Liu S R, Yang Y P. Studies on root biomass and productivity in dominant plantations population in the mountainous land in western Sichuan. Acta Ecologica Sinica, 2006, 26(2): 542-551. |
| [1] | 丁仲礼, 傅伯杰, 韩兴国, 葛全胜. 中国科学院“应对气候变化国际谈判的关键科学问题”项目群简介. 中国科学院院刊, 2009, 24(1): 8-17. |
| [2] | 李长生, 肖向明, Frolking S, Moore Ⅲ B, Salas W, 邱建军, 张宇, 庄亚辉, 王效科, 戴昭华, 刘纪远, 秦小光, 廖柏寒, Sass R. 中国农田的温室气体排放. 第四纪研究, 2003, 23(5): 493-503. |
| [4] | 陈佐忠. 中国典型草原生态系统. 北京: 科学出版社, 2000. |
| [12] | 王斌, 杨校生. 4种典型地带性森林生态系统碳含量与碳密度比较. 湖南农业大学学报: 自然科学版, 2010, 36(4): 465-469. |
| [14] | 彭晚霞, 王克林, 宋同清, 曾馥平, 王久荣. 喀斯特脆弱生态系统复合退化控制与重建模式. 生态学报, 2008, 28(2): 811-820. |
| [15] | 彭晚霞, 宋同清, 曾馥平, 王克林, 傅伟, 刘璐, 杜虎, 鹿士杨, 殷庆仓. 喀斯特常绿落叶阔叶混交林植物与土壤地形因子的耦合关系. 生态学报, 2010, 30(13): 3472-3481. |
| [16] | 宋同清, 彭晚霞, 曾馥平, 王克林, 覃文更, 谭卫宁, 刘璐, 杜虎, 鹿士杨. 木论喀斯特峰丛洼地森林群落空间格局及环境解释. 植物生态学报, 2010, 34(3): 298-308. |
| [17] | 宋同清, 彭晚霞, 曾馥平, 王克林, 曹洪麟, 李先琨, 覃文更, 谭卫宁, 刘璐. 喀斯特峰丛洼地不同类型森林群落的组成与生物多样性特征. 生物多样性, 2010, 18(4): 355-364. |
| [18] | 曾馥平, 彭晚霞, 宋同清, 王克林, 吴海勇, 宋希娟, 曾昭霞. 桂西北喀斯特人为干扰区植被自然恢复22年后群落特征. 生态学报, 2007, 27(12): 5110-5119. |
| [19] | 梁士楚. 贵阳喀斯特山地云贵鹅耳枥种群动态研究. 生态学报, 1992, 12(1): 53-60. |
| [20] | 宋敏, 彭晚霞, 邹冬生, 曾馥平, 杜虎, 鹿士杨. 喀斯特峰丛洼地不同森林表层土壤有机质的空间变异及成因. 生态学报, 2012, 32(19): 6259-6269. |
| [21] | 鲍士旦. 土壤农化分析. 北京: 农业出版社, 1988: 33-36. |
| [22] | 刘阳, 汪季, 刘静, 迟悦春, 张晓伟, 赵培林. 保护性耕作对玉米农田碳储量的影响. 内蒙古林业科技, 2012, 38(1): 14-17. |
| [23] | 李庆云. 黄淮海平原杨农间作系统碳贮量研究 [D]. 河南: 河南农业大学, 2008. |
| [24] | 吴征镒. 〈世界种子植物科的分布区类型系统〉的修订. 云南植物研究, 2003, 25(5): 535-538. |
| [25] | 吴征镒, 周浙昆, 李德铢, 彭华, 孙航. 世界种子植物科的分布区类型系统. 云南植物研究, 2003, 25(3): 245-257. |
| [26] | 何宁. 喀斯特峰丛洼地森林生态系统碳分配及其形成机制 [D]. 湖南: 中国科学院亚热带农业生态研究所, 2012. |
| [27] | 宫超, 汪思龙, 曾掌权, 邓仕坚, 陈建平, 龙康寿. 中亚热带常绿阔叶林不同演替阶段碳储量与格局特征. 生态学杂志, 2011, 30(9): 1935-1941. |
| [28] | 鲜骏仁, 张远彬, 王开运, 胡庭兴, 杨华. 川西亚高山5种森林生态系统的碳格局. 植物生态学报, 2009, 33(2): 283-290. |
| [29] | 韩美荣, 宋同清, 彭晚霞, 黄国勤, 杜虎, 鹿士杨, 时伟伟. 喀斯特峰丛洼地土壤矿物质的组成特征与作用. 应用生态学报, 2012, 23(3): 685-693. |
| [30] | 宋同清, 王克林, 彭晚霞, 王久荣, 肖润林, 曾馥平, 汤宇. 亚热带丘陵茶园间作白三叶草的生态效应. 生态学报, 2006, 26(11): 3647-3655. |
| [31] | 刘兴良, 马钦彦, 杨冬生, 史作民, 宿以明, 周世强, 刘世荣, 杨玉坡. 川西山地主要人工林种群根系生物量与生产力. 生态学报, 2006, 26(2): 542-551. |