 2014, Vol. 34
2014, Vol. 34文章信息
- 乌云娜, 雒文涛, 霍光伟, 李海山, 胡高娃
- WU Yunna, LUO Wentao, HUO Guangwei, LI Haishan, HU Gaowa
- 草原群落退化演替过程中微斑块土壤碳氮的空间异质动态
- Spatial heterogeneity dynamics of soil carbon and nitrogen in the mini-patches during degeneration succession of grassland communities
- 生态学报, 2014, 34(19): 5549-5557
- Acta Ecologica Sinica, 2014, 34(19): 5549-5557
- http://dx.doi.org/10.5846/stxb201301090067
-
文章历史
- 收稿日期:2013-1-9
- 网络出版日期:2014-3-7






2. 中国科学院沈阳应用生态研究所 森林与土壤生态国家重点实验室, 辽宁沈阳, 110016;
3. 中国科学院大学, 北京 100049;
4. 内蒙古呼伦贝尔市新巴尔虎右旗草原工作站, 内蒙古呼伦贝尔 021300
2. State Key Laboratory of Forest and Soil Ecology, Institute of Applied Ecology, Chinese Academy of Sciences, Shenyang 110164, China;
3. University of Chinese Academy of Sciences, Beijing 100049, China;
4. Environmental Bureau of Xinbaerhuyou County, Hulunber, Inner Mongolia 021300, China
生态学中的格局、过程、尺度和效应之间具有紧密的联系和相互作用。空间异质性反映了生态格局和生态过程的内在特性,是空间斑块性和空间梯度尺度依赖的综合反映。Li等[1]将空间异质性定义为所研究的系统特性在空间上的复杂性或变异性。土壤空间异质性是土壤的重要属性之一。土壤的形成过程包括物理、化学和生物过程,由于不同地区在气候、母岩、地形、植被和生物等方面的不同,形成了各种土壤类型,导致土壤性质存在明显的差异。在干旱、半干旱的草原生态系统中,土壤要素的小尺度空间异质性分布是一个普遍的特征[2, 3]。空间异质性一方面表现为地上植被空间格局的变化过程,另一方面表现为土壤资源变化过程,植被空间异质性和土壤资源的异质性是相互影响、相互制约的过程。植物群落结构的改变导致土壤养分的异质化,土壤养分的变化反过来会引起植被格局的变化[4, 5]。通常,生态系统在小尺度上表现出非平衡特征,而在大尺度上表现出复合稳定性特征。在放牧利用下,天然草地几乎无时不处于演替之中,而其中斑块的演替,特别是微斑块的变化无疑是最为活跃的成分。草原微斑块的形成与草地退化过程有着相同的推动力,斑块状植被相对稳定的空间格局是其适应环境的结果,也是草原植被存在的主要形式。草原植物群落的斑块格局与过程在小尺度上往往表现出更大的不稳定性,而且随着草地利用方式的扩展和程度的加剧,这种不稳定性将更趋剧烈。碳、氮是草原土壤生态系统中重要的养分要素,它们的空间变化影响着草原植被的发育与生长,因此揭示土壤碳、氮的空间异质性分布格局对于了解生态系统过程与动态具有重要意义。
呼伦贝尔草原是是我国目前景观生态类型和生物多样性较为丰富的草原生态系统类型,同时也是我国北方地区的重要生态防护屏障。但在长期的过牧与连续干旱的双重压力下,草原生态系统发生退化逆行演替,导致景观破碎化,土壤养分流失严重。目前,有关草地群落斑块化的报导大都集中于区域尺度上的研究,且对生态系统物质和能量输入、输出的研究也已经比较深入,但对草原群落微斑块尺度养分分布格局的报导却很少[6, 7]。鉴于以上认识,本文以呼伦贝尔草原克鲁伦河流域不同演替系列上的草原群落作为研究对象,在微生境尺度上,应用地统计分析方法研究不同演替过程中土壤全碳、全氮、碱解氮的空间分布格局与异质化程度,以期为进一步丰富“尺度-格局-过程”理论,阐明草原生态系统的退化机制提供科学依据。
1 材料与方法 1.1 研究区域概况研究区位于内蒙古呼伦贝尔草原克鲁伦河流域,地理位置47°36′—49°50′ N、115°31′—117°43′ E,气候属于温带半干旱季风气候,年降水量250—400 mm,湿润度0.5—0.7,年均温-3—0 ℃,全年≥10 ℃的活动积温1600—2000 ℃,潜在年蒸发量为950—1900 mm,土壤类型主要为栗钙土。植物种类主要有克氏针茅(Stipa krylovii)、羊草(Leymus chinensis)、糙隐子草(Cleistogenes squarrosa)、黄囊苔草(Carex korshinskyi)、冷蒿(Artemisia frigid)、银灰旋花(Convolvulus ammannii)、兴安天门冬(Asparagus dahuricus)、细叶葱(Allium tenuissimum)、野韭(Allium ramosum)、多根葱(Allium polyrhizum)等。
1.2 实验样地设置依据群落优势种的变化,确定放牧压力下的3个退化演替系列分别为:克氏针茅+羊草群落、克氏针茅+糙隐子草群落、多根葱+黄囊苔草群落3个演替序列。结合当地畜牧局提供的相关数据,计算出3个样地的载畜量分别为0.62 羊/hm2、1.55 羊/hm2、2.79 羊/hm2。在此分别定义为演替前期、演替中期、演替后期3个退化演替序列。
演替前期样地(N 49° 14′ 71.1″ E 116° 55′ 59.0″)围栏封育时间为2001年,为季节性放牧,植物在生长季不受人为干扰;演替中期样地(N 47° 55′ 07.6″ E 117° 23′ 40.6″)为年季间轮流放牧区,植物能够进行间歇性恢复生长;演替后期样地(N 48° 31′91.4″ E 116° 40′ 19.7″)为自由放牧区,常年受到放牧压力影响,植物生长常年受到人为干扰。3个样地地形平坦、海拔相似,土壤类型均为栗钙土。
1.3 实验设计与测定方法于2011年植物生长季(7月中旬),在退化演替系列样地内,选择微地形条件一致且地势平坦、植被均匀的典型地段,设置面积为1 m×1 m的3个样方,采用均匀网格法,按10 cm×10 cm的密度把每个样方平均分为100个小格子(图 1),对每个小格子进行编号。采用刈割法采集每个小格子内的所有植物,在每个小格子中心部位用100 cm3环刀取土(0—10 cm),装袋密封并带回实验室。土壤样品经自然风干后剔除植物根系等杂质,并用球磨仪磨碎过1 mm土壤筛,采用重铬酸钾容量法-外加热法测定土壤全碳含量,采用半微量凯氏定氮法测定土壤全氮,采用碱解扩散法测定土壤碱解氮[8]。

|
| 图 1 样方设定示意图 Fig. 1 Schematic diagram of sampling design with three sites |
用SPSS13.0软件对不同样地的全碳、全氮、碱解氮进行差异度(方差分析)和正态分布检验。全碳、全氮、碱解氮数据都进行了对数(log)转化使其符合正态分布,以消除变异性对回归显著性的影响。采用地统计学的分析软件(GS+ 9.0和Surfer 8.0软件)分析土壤全碳、全氮、碱解氮的空间异质性。
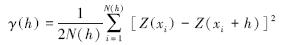
式中,γ(h)为半方差函数,h为两样本间的分离距离,Z(xi)和Z(xi+h)分别为随机变量Z在空间位置xi和xi+h上的取值,N(h)为在空间上具有相同间隔距离h的样本对总数。块金值(nugget)是间隔距离h→0时,γ(0)的极限值。随着间隔距离h的增加变异函数γ(h)值从块金值(nugget)变为一个相对稳定的常数基台值(sill)。块金系数(N ∶ S值)是块金方差与基台值之比。它表示(空间变异性程度)由随机性因素引起的空间异质性占系统总变异的比例。该比值高,说明由随机部分引起的空间变异性程度较大;相反则由结构性因素引起的空间变异性程度较大,如果该比值接近1,则说明该变量在整个尺度上具有恒定的变异。
2 结果与分析 2.1 退化演替系列上土壤养分元素描述性特征由表 1可知,草原演替过程中草原群落植被土壤全碳、全氮、碱解氮分布都发生了明显变化(P<0.05)。以10 cm×10 cm的样方为基本单元进行统计,演替前期样地土壤全碳含量平均值为43.63 g/kg,变化范围为32.78—65.47 g/kg;演替中期样地平均值为24.47 g/kg,变化范围为13.05—54.86 g/kg;演替后期样地平均值为31.85 g/kg,变化范围为20.83—40.43 g/kg。演替前期、演替中期、演替后期样地土壤全氮含量变化范围分别为2.03—5.00 mg/kg、1.11—1.87 mg/kg、1.32—5.54 mg/kg,土壤全氮含量表现为演替前期(2.56 g/kg)>演替后期(1.92 g/kg)>演替中期(1.40 g/kg)(P<0.05)。土壤碱解氮含量在演替前期样地上变化范围为103.35—293.45 g/kg,平均值为174.46 g/kg,演替中期样地上为62.75—280.53 mg/kg,平均值为117.69 mg/kg,演替后期样地上为59.06—287.92 mg/kg,平均值为131.49 mg/kg。不同演替群落土壤全碳、全氮、碱解氮含量变化为演替前期>演替后期>演替中期(P<0.05)。
| 变量 Variable | 样地 Sites | 最小值 Minimum | 最大值 Maximum | 均值 Mean | 标准差 Standard deviation | 变异系数 Variation coefficient |
| 不同字母表示处理间差异显著(P<0.05) | ||||||
| 全碳/(g/kg) | 演替前期 | 32.78 | 65.47 | 43.63a | 5.72 | 0.15 |
| Soil total carbon | 演替中期 | 13.05 | 54.86 | 24.17c | 5.70 | 0.24 |
| 演替后期 | 20.83 | 40.43 | 31.85b | 4.10 | 0.13 | |
| 全氮/(g/kg) | 演替前期 | 2.03 | 5.00 | 2.56a | 0.47 | 0.18 |
| Soil total nitrogen | 演替中期 | 1.11 | 1.87 | 1.40c | 0.15 | 0.11 |
| 演替后期 | 1.32 | 5.54 | 1.92b | 0.45 | 0.23 | |
| 碱解氮/(mg/kg) | 演替前期 | 103.35 | 293.45 | 174.46a | 30.94 | 0.18 |
| Alkali-hydrolyzable | 演替中期 | 62.75 | 280.53 | 117.69c | 38.57 | 0.33 |
| nitrogen | 演替后期 | 59.06 | 287.92 | 131.49b | 34.01 | 0.26 |
变异系数(CV)反映了总体的内部变异程度。通常认为,变异系数CV≤0.1时为弱变异性;0.1<CV<1时为中等变异性;CV≥1时为强变异性[11]。在土壤全碳含量的变异性分析中,演替前期、中期和后期样地的变异系数分别为0.13、0.24、0.13,均属于中等变异,在演替中期样地上变异系数最大。从土壤全氮含量变异来看,演替前期、演替中期、演替后期样地的变异系数分别为0.18、0.11、0.23,均属于中等变异,变异系数表现为先减小再增大的趋势,演替后期样地上变异程度最大。土壤碱解氮含量变异系数为演替前期(0.18)<演替后期(0.26)<演替中期(0.33)。
2.2 退化演替过程中土壤营养元素的空间分异性基台值(Still)表示系统内总的变异,块金值与基台值的比值称为块金系数(N ∶ S),按照变量空间相关性程度的分级标准,块金系数<25%表明变量之间具有强烈的空间相关性,说明在影响其分布空间变异的因素中,结构性因素占据绝对的主导地位;块金系数25%—75%表明变量具有中等空间自相关,其变异是由结构性因素和随机性因素共同作用的结果;块金系数>75%时,变量空间自相关性微弱,随机变异占主导因素。从表 2可以看出,土壤全碳含量变异函数块金系数随退化演替程度的增加表现出先减小再增大的趋势,空间自相关性先变大然后变小。土壤全碳在演替后期样地上表现出中等程度的空间自相关,而在演替中期和演替前期样地上表现为强烈空间自相关。土壤全氮含量变异函数块金系数表现为演替中期>演替前期>演替后期,空间自相关性为演替后期>演替前期>演替中期,在3个放牧样地上均呈现出显著的空间相关性。土壤碱解氮含量在演替前期样地上属于中等程度的空间自相关,而在演替中期和轻演替后期样样地上均表现为明显的空间自相关性。
| 变量 Variable | 样地 Sites | 理论模型 Model | 块金值 Nugget | 拱高 C1 | 基台值 Still | 块金系数 N ∶ S |
| 全碳 | 演替前期 | 球状 | 1.7 | 11.23 | 12.93 | 13.1 |
| Soil total carbon | 演替中期 | 球状 | 0.4 | 15.47 | 15.88 | 2.6 |
| 演替后期 | 球状 | 9.9 | 26.28 | 36.19 | 27.4 | |
| 全氮 | 演替前期 | 球状 | 0.4 | 0.12 | 0.239 | 16.6 |
| Soil total nitrogen | 演替中期 | 球状 | 0.5 | 0.15 | 0.191 | 23.0 |
| 演替后期 | 球状 | 0.6 | 0.02 | 0.021 | 2.4 | |
| 碱解氮 | 演替前期 | 球状 | 197 | 1493 | 2690 | 44.5 |
| Alkali-hydrolyzable | 演替中期 | 球状 | 11 | 1034 | 1045 | 1.1 |
| nitrogen | 演替后期 | 球状 | 24 | 1493 | 1517 | 1.6 |
为了更加直观地反映土壤元素空间水平的分布特征,采用Surfer软件绘制了土壤全碳、全氮、碱解氮的等值线图(图 2)。图中,斑块颜色从浅到深(即白色到黑色)表示土壤中养分含量由多到少的变化规律。可以看出,(a)演替前期样地上主要是白色斑点和灰白色斑点,土壤全碳含量很高但异质性不高。演替中期阶段上主要是黑色斑点和白灰色斑点,全碳含量很小但异质性很高。演替后期样地地上主要是灰白色斑点,全碳含量居中,异质性不高。(b)演替前期样地主要以灰白色斑块为主,土壤全氮含量很高,异质性中等。演替中期样地上出现了一个异常极大值,去除此点后主要以黑色和黑灰色斑点为主,全氮含量很低,但异质性很大。演替后期样地上主要以灰白色斑点为主但在局域出现了白色斑点,全氮含量居中,异质性很小。(c)演替前期样地主要是以灰白色和白色斑点为主,碱解氮含量很高,异质性很小。演替中期样地主要是以灰白色,黑灰色和极小局部出现的白色为主,碱解氮含量很低,异质性很大。演替后期样地主要是以灰白色和少许白色、黑色斑点,碱解氮含量居中,异质性中等。

|
| 图 2 演替过程中土壤全碳、全氮和碱解氮的等值线图 Fig. 2 Counter map of soil total carbon,total nitrogen and alkali hydrolyzable nitrogen under succession degradation |
在内蒙古地区,由于气候变化和人为干扰,潜在草地类型之间发生了复杂的演替过程。温带草原演替过程中,植物种类、生活型、根系类型发生了变化,强烈影响到生态系统的进化过程。草原演替系列是一个以低能量水平进行自我调控自我维持的草原生态系统,具有一定的稳定性。蒙古高原东部克氏针茅草原在连续多年的干旱和放牧压力下退化明显,退化程度逐年加重,群落中的优势种和伴生种发生了显著变化,退化演替序列为克氏针茅+羊草群落、克氏针茅+糙隐子草群落、多根葱+黄囊苔草群落3个序列。从群落种类成分分析来看,糙隐子草(C4植物)与克氏针茅(C3植物)相比,具有低呼吸、低CO2补偿点、高光饱和点、高的净光合速率和光量子效率等特点,同是对水分、氮素、光及CO2利用效率均高于克氏针茅(C3植物),糙隐子草(C4植物)比克氏针茅(C3植物)具有生理优势。近年来该地区温度持续升高[12],C3植物逐渐的被更具有生理优势的C4植物所取代[13]。Wang等[14]认为温度是控制C3与C4植物分布的主要因子,随温度的升高C4植物的竞争力会更强。多根葱具有极强的吸收能力,根系粗壮发达并向周围扩展,能够迅速吸收土壤表层水分,另外,多根葱在水分适宜的条件下瞬间生长速率远远大于糙隐子草植物,具有非常强的储水能力,含水量高达80%。这些特点多根葱在重度退化系列上竞争排斥其他物种(糙隐子草),并迅速构建优势群落创造了条件。
放牧活动是内蒙古呼伦贝尔克鲁伦河流域天然草原土地利用的主要方式,长期放牧导致草原生态系统的退化演替。退化草原中植物通过对土壤全碳、全氮和碱解氮的吸收和沉积,将其集中于其冠层下土壤中,从而引起土壤资源的空间异质性[15, 16, 17]。本研究结果表明:土壤全碳、全氮和碱解氮含量均表现为演替前期>演替后期>演替中期(P<0.05)。这一结果可以由全碳含量、全氮和碱解氮3个变量之间的显著相关关系来进行解释(P<0.001)(图 3),全碳与全氮、全碳与碱解氮、全氮与碱解氮之间的解释度分别为77%,37%,42%。Tian等通过中国2473组土壤碳、氮元素数据进行分析,同样发现土壤表层全碳和全氮元素含量之间呈极显著正相关性(R2=0.82,P<0.001)[18]。从土壤全碳、全氮和碱解氮的变异系数和变异函数综合分析来看,10 cm×10 cm微尺度上草原退化演替中土壤全碳、全氮和碱解氮的空间异质性具有明显的不一致性。斑块是基质植被在受到局部强扰动下逆行演替的结果,具有高度的不稳定性,同时还反映出草地退化过程的非均质性特点。全碳的空间异质性表现为演替中期>演替前期>演替后期,全氮表现为演替后期>演替前期>演替中期,碱解氮表现为演替中期>演替后期>演替前期。这一结果说明,当草原群落达到中度退化阶段时,放牧干扰对草地的影响已经达到由量变到质变的临界区,如不及时降低放牧压力,草地的生态和经济性质将发生根本性的转变。土壤养分斑块的产生与动态受控于自然的和人为的干扰作用,干扰是破坏生态系统、群落或种群结构,以及改变资源和基质可利用性或自然环境的任何在时间上相对不连续的事件,这是驱动异质性发生的主要来源。干扰因素在空间和时间上往往是杂乱无章的,使斑块的形成 发生在不同的时间和地点,从而形成一种动态的镶

|
| 图 3 演替过程中土壤全碳、全氮和碱解氮之间的相互关系 Fig. 3 The relationships between soil total carbon (C) and soil total nitrogen (N) ,between soil total carbon (C) and alkali hydrolyzable nitrogen (AN) and between soil total nitrogen (N) and alkali hydrolyzable nitrogen (AN) across the three succession degradation sites |
嵌体结构[19, 20, 21]。Zuo等[22]研究表明,我国科尔沁沙地的土壤性质的空间分布格局与沙丘土壤颗粒粒度、采样点的相对高度、植被覆盖、土壤有机碳以及总氮密切相关(P<0.05)。Zheng等[23]对我国贺兰山地区的相关研究表明,猪毛菜属对土壤理化性质(水分、有机质、全氮及酸碱度)具有重要的作用,并建立了草原退化演替系列的概念模型。
小尺度上植物群落退化系列的空间异质性研究,可以加深对植物群落与土壤相互作用过程与耦合机制的理解,可以深入地了解土壤元素在草原斑块化过程中的影响和作用。近年来草原灌丛化过程机制、草原灌丛化的生物地化循环后果以及它对气候变化的影响日趋受到国内外生态学家的重视,已经成为陆地生态系统全球变化研究的重要领域。金钊等[24]研究表明草原灌丛化过程中草地生态系统土壤养分及相关元素在水平和垂直方向发生分异,碳、氮生物元素地球化学循环变化将对全球气候变化产生显著作用。草原微斑块化动态不仅存在于草原灌丛化过程中,而是普遍存在于草原的任何动态过程中。斑块化格局与植物本身的生长和进化特性密切关系,在长期的进化与适应过程中,植物趋向于拥有较大面积的枝叶群从而可以更加有效的进行植物光合效率,同时对空中的碳、氮等营养元素进行有效截留。同时,进化形成的庞大植物根系组织群可以更有效吸收土壤营养元素。植物枝叶群、根系群集中起来的元素,不断的循环于植物-土壤这个功能有机体中,通过改变土壤结构、微生物生物量、土壤湿度和小气候,并将营养元素集中于其植冠层之下,从而对养分的空间分布和循环产生影响,使土壤资源异质性增强。反过来这种异质性又会加速植被的空间格局分异,从而导致“养分聚集效应”的形成[25, 26, 27, 28, 29, 30]。养分聚集是土壤资源异质性分布的高度表现方式,植物枝叶群和根系群占用空间相对越大,效应相对越强。“养分聚集效应”的强弱程度是草原荒漠化的一个重要指标,荒漠化或草地退化过程中的大量研究支持这一概念的生物生态学解释。例如,Li等[31]认为干旱、半干旱地区的草地草本植物为优势的群落被灌丛群落所替代是草地发生退化或荒漠化的显著特征,该过程增加草地土壤资源的空间分布异质性,使土壤-植被系统的生物过程愈来愈多的集中在植冠下的养分聚集范围内。“养分聚集效应”概念的提出,将进一步丰富“尺度-格局-过程”理论,推动草原生态系统的机理研究。
| [1] | Li H, Reynolds J F. On definition and quantification of heterogeneity. Oikos, 1995, 73(2): 280-284. |
| [2] | Chen Y F, Dong M. Spatial heterogeneity in ecological systems. Acta Ecologica Sinica, 2003, 23(2): 346-351. |
| [3] | Webster R. Quantitative spatial analysis of soil in the field. Advance in Soil Science, 1985, 3: 1-70. |
| [4] | Wang C Q, Wang Q C. The spatial heterogeneity of soil physical properties in forests. Acta Ecologica Sinica, 2000, 20(6): 945-950. |
| [5] | Peco B, Sánchez A M, Azcárate F M. Abandonment in grazing systems: Consequences for vegetation and soil. Agriculture, Ecosystems & Environment, 2006, 113(1/4): 284-294. |
| [6] | Zhang W G, Hang W B, Yang Z Y. The study on the relationship between mini-patch and degradation of pasture. Acta Prataculturae Sinica, 2003, 12(3): 44-50. |
| [7] | Bai Y F, Xu Z X, Li D X. On the small scale spatial heterogeneity of soil moisture, carbon and nitrogen in Stipa communities of the Inner Mongolia Plateau. Acta Ecologica Sinica, 2002, 22(8): 1210-1218. |
| [8] | Jackson ML. Soil chemical analysis. Englewood Cliffs, New Jersey: Prantice Hall Inc.; 1958. |
| [9] | Schlesinger W H, Raikes J A, Hartley A E, Cross A F. On the spatial pattern of soil nutrients in desert ecosystems. Ecology, 1996, 77(2): 364-374. |
| [10] | William H S, Jane A R, Anne E H, Anne F C. On the Spatial Pattern of Soil Nutrients in Desert Ecosystems. Ecology, 1996, 77:364-374. |
| [11] | Cambardella C A, Moorman T B, Parkin T B, Karlen D L, Novak J M, Turco R F, Konopka A E. Field-scale variability of soil properties in central Iowa soils. Soil Science Society of America Journal, 1994, 58(5): 1501-1511. |
| [12] | Wu Y N, Luo W T, Hao P, Shiyomi M, Lv J Z, Huo G W. Climate change and the effect on the biomass growth in steppe communities in Kherlon River Basin, Northern China. Acta Ecologica Sinica, 2012, 32(4): 215-220. |
| [13] | Pau S, Edwards E J, Still C J. Improving our understanding of environmental controls on the distribution of C3 and C4 grasses. Global Change Biology, 2013, 19(1): 184-196. |
| [14] | Wang R Z. C4 species and their response to large-scale longitudinal climate variables along the Northeast China Transect (NECT). Photosynthetica, 2004, 42(1): 71-79. |
| [15] | Sparrow A D. A heterogeneity of heterogeneities. Trends in Ecology & Evolution, 1999, 14(11): 422-423. |
| [16] | Xiong X G, Han X G. Spatial heterogeneity in soil carbon and nitrogen resources, caused by Caragana microphylla, in the thicketization of semiarid grassland, Inner Mongolia. Acta Ecologica Sinica, 2005, 25(7): 1673-1683. |
| [17] | Xiong X G, Han X G, Bao Y J. Discussion on the research into sandy desertification, accompanying by thicketization of semiarid grasslands in Inner Mongolia, China. Acta Prataculturae Sinica, 2005, 14(5): 1-5. |
| [18] | Tian H Q, Chen G S, Zhang C, Melillo J M, Hall C A S. Pattern and variation of C: N: P ratios in China's soils: a synthesis of observational data. Biogeochemistry, 2009, 98(1/3): 139-151. |
| [19] | Thompson K. Small-scale heterogeneity in seed bank of an acidic grassland. The Journal of Ecology, 1986, 74(3): 733-738. |
| [20] | Luck M, Wu J G. A gradient analysis of urban landscape pattern: a case study from the Phoenix metropolitan region, Arizona, USA Landscape Ecology, 2002, 17: 327-339. |
| [21] | Xiong X G, Han X G, Chen Q S. The walter island effect in the arid and semi-arid ecosystems//Li C S, ed. Advances in plant science. Beijing: Higher Education Press, 2003. |
| [22] | Zuo X A, Zhao H L, Zhao X Y, Zhang T H, Guo Y R, Wang S K, Drake S. Spatial pattern and heterogeneity of soil properties in sand dunes under grazing and restoration in Horqin Sandy Land, Northern China. Soil & Tillage Research, 2007, 99(2): 202-212. |
| [23] | Zheng J, He M, Li X, Chen Y, Li X, Liu L. Effects of Salsola passerina shrub patches on the microscale heterogeneity of soil in a montane grassland, China. Journal of Arid Environments, 2008, 72(3): 150-161. |
| [24] | Jin Z, Yu C, Dong Y S. Shrub encroachment and accompanied changes of biogeochemistry cycles in semi-arid and arid grasslands. Progress in Geography, 2007, 26(4): 23-32. |
| [25] | Robertson G P. Geostatistics in ecology: interpolating with known variance. Ecology, 1987, 68(3): 744-748. |
| [26] | Chen F S, Zeng D H. Spatial structure of soil nutrients of degraded grassland in southeastern Keerqin sand land and its ecological explanations. Acta Prataculturae Sinica, 2004, 13(1): 39-44. |
| [27] | Xiong X G, Han X G. New field of ecology-fertile island effects and thicketization. Plant Journal, 2001, (3): 45-46. |
| [28] | Zhang H, Shi P J, Zheng H Q. Research progress in relationship between shrub invasion and soil heterogeneity in natural semi-arid grassland. Acta Phytoecologica Sinica, 2001, 25(3): 366-370. |
| [29] | Whitehead D C. Nutrient Elements in Grassland: Soil-Plant-Animal Relationships. Wallingford, UK: CABI Publishing, 2000. |
| [30] | Kubien D S, Sage R F. Low-temperature photosynthetic performance of a C4 grass and a co-occurring C3 grass native to high latitudes. Plant, Cell & Environment, 2004, 27: 907-916. |
| [31] | Li X R, Wang X P, Li T, Zhang J G. Microbiotic soil crust and its effect on vegetation and habitat on artificially stabilized desert dunes in Tengger desert, North China. Biology and Fertility of Soils, 2002, 35(3): 147-154. |
| [2] | 陈玉福, 董鸣. 生态学系统的空间异质性. 生态学报, 2003, 23(2): 346-351. |
| [4] | 王政权, 王庆成. 森林土壤物理性质的空间异质性研究. 生态学报, 2000, 20(6): 945-950. |
| [6] | 张卫国, 黄文冰, 杨振宇. 草地微斑块与草地退化关系的研究. 草业学报, 2003, 12(3): 44-50. |
| [7] | 白永飞, 许志信, 李德新. 内蒙古高原针茅草原群落土壤水分和碳、氮分布的小尺度空间异质性. 生态学报, 2002, 22(8): 1210-1218. |
| [16] | 熊小刚, 韩兴国. 内蒙古半干旱草原灌丛化过程中小叶锦鸡儿引起的土壤碳、氮资源空间异质性分布. 生态学报, 2005, 25(7): 1673-1683. |
| [17] | 熊小刚, 韩兴国, 鲍雅静. 试论我国内蒙古半干旱草原灌丛沙漠化的研究. 草业学报, 2005, 14(5): 1-5. |
| [21] | 熊小刚, 韩兴国, 陈全胜. 干旱和半干旱生态系统中的沃岛效应 // 李承森. 植物科学进展(5). 北京: 高等教育出版社, 2003. |
| [24] | 金钊, 玉春, 董云社. 干旱半干旱地区草原灌丛荒漠化及其生物地球化学循环. 地理科学进展, 2007, 26(4): 23-32. |
| [26] | 陈伏生, 曾德慧. 科尔沁沙地退化草场土壤养分的空间结构分析. 草业学报, 2004, 13(1): 39-44. |
| [27] | 熊小刚, 韩兴国. 生态学中的新领域——沃岛效应与草原灌丛化. 植物杂志, 2003, (3): 45-46. |
| [28] | 张宏, 史培军, 郑秋红. 半干旱地区天然草地灌丛化与土壤异质性关系研究进展. 植物生态学报, 2001, 25(3): 366-370. |