 2014, Vol. 34
2014, Vol. 34文章信息
- 黄志敏, 陈椽, 刘之威, 龙胜兴
- HUANG Zhimin, CHEN Chuan, LIU Zhiwei, LONG Shengxing
- 贵州百花湖夏季浮游植物昼夜垂直分布特征
- Diel vertical distribution patterns of phytoplanktion in Baihua Reservoir, Guizhou Province
- 生态学报, 2014, 34(19): 5389-5397
- Acta Ecologica Sinica, 2014, 34(19): 5389-5397
- http://dx.doi.org/10.5846/stxb201301070054
-
文章历史
- 收稿日期:2013-1-7
- 网络出版日期:2014-3-7






2. 贵阳市环境监测中心站, 贵阳 550002
2. Environmental Monitoring Centre of Guiyang City, Guiyang 550002, China
浮游植物(phytoplankton) 作为水生态系统的初级生产者和食物链的第一环节,对维持整个水生态系统稳定性和完整性起到重要的作用[1],其群落结构及其变化情况是水体水质的一个重要表征指标[2]。近年来,藻类水华的形成是湖泊富营养化的重要标志,其危害备受关注。关于水华的爆发机制及其治理的研究还有欠缺,而引起水华藻类群体迁移和聚集的机制是这方面研究的重要内容[3]。无论在海洋和淡水水体中,浮游植物在水柱中垂直分布格局昼夜变化的现象比较广泛[4],尤其是鞭毛藻[5, 6, 7]和具有伪空泡的蓝藻[8, 9],鞭毛藻昼夜垂直迁移是其生存能力的体现,白天垂直向上迁移吸收更多的光进行光合作用,晚上向下迁移吸收营养[10]。具有伪空泡的蓝藻具有浮力和随着环境条件的改变而对浮力进行调节的能力[11, 12, 13],从而在水体中占据优势地位。光照强度和温度对浮游植物昼夜分布和垂直迁移的影响较大。Spencer 等认为光照条件与藻类在水体中聚集、漂移乃至水华形成有着密切关系[14]。孟氏浮游蓝藻丝在水体中的垂直变化对光照的响应明显[15]。失去浮力的越冬蓝藻在20℃能够较快恢复浮力,而在12℃时则恢复较慢[16]。具有昼夜垂直迁移能力的藻类在水体中具有竞争优势,在适宜的条件下可能导致水华的发生,致使水质恶化。迄今为止,关于贵州喀斯特深水水库浮游植物昼夜垂直分布的研究尚未见报道。依据李秋华、胡晓红等报道:近年来夏季百花湖浮游植物群落结构已发生明显变化[17, 18, 19]。本文以贵州百花湖为研究对象,对浮游植物的垂直分布和迁移进行研究,以期揭示浮游植物在水体中的垂直分布格局及迁移规律,确定影响浮游植物垂直分布和迁移的主要环境因子,为贵州喀斯特深水水库水华爆发机制研究积累基础数据。
1 材料与方法 1.1 样点设置和采样时间百花湖位于贵阳西北郊,是猫跳河梯级电站第二级蓄水库。主要由红枫湖下泄水以及长冲河、东门桥河、麦城河和麦西河四源汇成。采样点S1位于百花湖库心(图 1),于2012年7月31日—8月1日白天8:00、10:00、12:00、14:00、16:00、18:00和夜晚20:00、22:00、2:00、6:00进行昼夜24h定点分层(0.5、1、2、3、4、5、6、7、8、9、10、11、12、13、14m)监测。监测指标为水温(WT)、光合有效辐射(PAR)、浮游植物丰度。

|
| 图 1 百花湖样点设置图 Fig. 1 Sampling sites in Baihua Reservoir |
表层浮游植物定性样品使用25号浮游生物网在0.5 m处呈“∞”字形缓慢捞取15 min,1—14m浮游植物定性样品使用采水器分别在各水层采水10升,用25号浮游生物网过滤,均取30 mL,加体积比为甘油 ∶ 甲醛 ∶ 水=1 ∶ 1 ∶ 8的固定液固定。浮游植物定量样品,取水1000 mL,加入15 mL鲁哥试剂固定,带回实验室静置沉淀48 h,取30 mL底层浓缩液。浮游植物定性、定量样品均在显微镜下进行鉴定和计数。计数方法采用目镜视野法,视野数为100个[20, 21, 22]。光照强度采用ZDS-10W水下照度计测量,水温使用YSI便携式水质多参数测定仪现场测定。
1.3 数据分析实验所得数据用Excel进行分析处理,用SPSS18.0进行相关性分析。浮游植物丰度、湖泊假鱼腥藻丰度的昼夜垂直分布格局采用时间深度等值图进行模拟。时间深度等值图在Surfer软件(Golden software surfer 8.0) 中绘制,绘制过程中选用Kriging插值模拟。同时选用Morisita指数(morisita′s index,MI) 进一步定量检验浮游植物在水体中的垂直分布格局,该指数的计算公式为[23]:
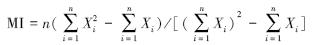
式中,n为水柱取样的分层数,x为水柱中各层的浮游植物丰度。当MI=1 时,表示浮游植物在水柱中随机分布(random distribution) ;当MI<1 时,表示浮游植物在水柱中均匀分布 ;当MI>1 时,表示浮游植物在水柱中集群分布[24, 25, 26]。 将Morisita 指数与入射光光照强度、水柱温差(WCTD) 进行Spearman相关分析,用以检验光照、水温对浮游植物分布格局昼夜变化的影响。
2 结果 2.1 浮游植物种类组成及垂直分布2012年7月31日—8月1日,百花湖水库共检出浮游植物151种(含变种和变型),隶属于7门9纲16目30科57属。其中绿藻门(Chlorophyta)种类最多,为28属87种,约占总种数的57.62%;蓝藻门(Byanobacteria)11属21种,约占总种数的13.91%;硅藻门(Bacillariophyta)12属31种,约占总种数的20.53%;隐藻门(Cryptophyta)2属5种,约占总种数的3.31%;甲藻门(Pyrrophyta)4属3种,约占总种数的1.99%,金藻门(Chrysophyta)1属2种,约占总种数的1.33%,裸藻门(Euglenophyta1)1属2种,约占总种数的1.33%。
从图 2可以看出: 各水层均以绿藻种类数最多,4 m以浅(除3 m外)处蓝藻种类数大于硅藻,4 m以深处绿藻>硅藻>蓝藻。蓝藻、绿藻、硅藻种类数在水体中具有明显的分层现象。蓝藻种类数在0.5—2 m较多,2 m以深水层垂直变化不明显;绿藻种类数在1 m处出现峰值,总体表现为0.5—6 m较多,7—14 m较少;硅藻种类数在0.5—5 m较少,从6 m水深处开始明显增多。浮游植物总种类数垂直分布波动较大,0.5—2 m浮游植物总种类数较多,与蓝藻、绿藻表现一致,6 m处硅藻种类数明显增多,导致浮游植物总种类数明显高于邻近水层,7 m及以深水层,浮游植物总种类数呈上升趋势,由硅藻种类增加数大于绿藻种类减少数决定。其它藻门种类数较少,且垂直分布变化不明显。

|
| 图 2 百花湖浮游植物种类数垂直变化趋势 Fig. 2 The vertical distribution about number of phytoplankton species in Baihua Reservoir |
以物种丰度的5%作为优势属依据[27],湖泊假鱼腥藻(Pseudanabaena limnetica)在各水层均占优势。此外,蓝藻门的小尖头藻属(Raphidiopsis Fritsch et Rich.)、颤藻属(Oscillatoria Vauch. ex Gom.)、 微囊藻属(Microcystis Kütz.),隐藻门的隐藻属(Cryptomonas Ehr.)、蓝隐藻属(Chroomonas Hangsg.),绿藻门的空球藻(Eudorina elegans Ehr.)分别在部分水层表现出优势地位,无明显的垂直变化,对浮游植物总体垂直分布格局影响不大。
2.2 浮游植物丰度昼夜垂直分布从图 3可以看出,各水层浮游植物丰度最高达73.85×106个/L,出现在12:00时的2 m水深处,平均丰度为18.06×106个/L,主要聚集在水体的0.5—4 m。其中8:00时浮游植物在2—3 m处聚集程度最高,10:00时主要聚集在0.5 m处,10:00—12:00时浮游植物的细胞丰度呈由0.5 m向2 m扩增的趋势,12:00时在2 m处达到峰值,12:00—14:00呈由2 m向0.5 m扩增的趋势,16:00时又大量聚集于1 m处。夜间浮游植物的聚集程度变化幅度不明显。此外,图 3还表明:水体中浮游植物总丰度夜间低于白天。

|
| 图 3 百花湖浮游植物总丰度的时间深度等值图 Fig. 3 Depth-time distribution of densities of total phytoplankton in Baihua Reservoir |
从图 4可以看出,湖泊假鱼腥藻为绝对优势种,相对丰度达82.69%,平均丰度为14.93×106个/L,主要聚集在水体的0.5—4 m。其中8:00时湖泊假鱼腥藻在2—3 m处聚集程度最高,10:00时主要聚集在0.5 m处,10:00—12:00时湖泊假鱼腥藻的细胞丰度呈由0.5 m向2 m扩增的趋势,12:00时在2 m处达到峰值,为62.31×106个/L,此时0.5 m处的细胞丰度是除6:00外的最小值,为25.41×106个/L,12:00—14:00呈由2 m向0.5 m扩增的趋势,16:00时又大量聚集于1 m处。可见,湖泊假鱼腥藻在白天具有明显的垂直迁移现象。夜间湖泊假鱼腥藻的聚集程度变化幅度不明显。湖泊假鱼腥藻的时间深度等值图的变化趋势与浮游植物总丰度一致。因此水体中湖泊假鱼腥藻细胞丰度的昼夜垂直变化规律能够反映浮游植物总丰度的变化规律。此外,图 4还表明:水体中湖泊假鱼腥藻丰度表现为夜间低于白天。除湖泊假鱼腥藻外其他各种类在垂直断面上呈不连续分布或密度极低。

|
| 图 4 百花湖湖泊假鱼腥藻丰度的时间深度等值图 Fig. 4 Depth-time distribution of densities Pseudanabaena limnetica in Baihua Reservoir |
从图 5可以看出,浮游植物总丰度的MI指数在一昼夜时间内均大于1,说明浮游植物在水体中昼夜均呈聚集分布,浮游植物总丰度的MI指数白天在1.45—1.93之间,夜间在1.31—1.42之间,说明白天浮游植物在水体中聚集程度高于夜间。湖泊假鱼腥藻丰度的MI指数在一昼夜时间内均大于1,说明湖泊假鱼腥藻在水体中昼夜均呈聚集分布,湖泊假鱼腥藻丰度的MI指数白天在1.45—2.07之间,夜间在1.40—1.46之间,说明白天湖泊假鱼腥藻在水体中聚集程度高于夜间。图 5还表明,白天浮游植物总丰度和湖泊假鱼腥藻丰度聚集程度的变化幅度波动较大,夜间较平缓,湖泊假鱼腥藻垂直分布格局的昼夜变化趋势与浮游植物总丰度一致。

|
| 图 5 百花湖浮游植物总丰度和湖泊假鱼腥藻丰度的MI指数昼夜变化趋势 Fig. 5 Morisita′s index (MI) of total phytoplankton densities and Pseudanabaena limnetica densities in Baihua Reservoir |
如图 6,百花湖水库夏季水下0.5—2 m的光照强度随时间的变化与太阳光强度变化基本一致。从早晨到12:00逐渐增强,12:00时达到最大值,分别为219、189、42 μmol m-2 s-1,12:00以后逐渐减弱。14:00时,2 m以深至5 m处光照强度高于其它时间的相同水层,可能是天气原因,或水体透明的影响,5 m以深处各时间的光照强度均小于0.5 m处光照强度的1%。各时间水体0.5—2 m光照强度的垂直变化幅度存在差异。10:00、l2:00、14:00时的变化幅度较大,分别在18—121 μmol m-2 s-1、42—219 μmol m-2 s-1、40—148 μmol m-2 s-1之间,12:00时最大。8:00、16:00、18:00时变化幅度较小,分别在17—54 μmol m-2 s-1、23—28 μmol m-2 s-1、6—24 μmol m-2 s-1 之间,16:00时最小。2 m以深水层光照强度的垂直变化幅度不大。

|
| 图 6 百花湖水库水下光合有效辐射的时间深度等值图 Fig. 6 Depth-time distribution of photosynthetically available radiation in Baihua Reservoir |
如图 7,调查期间百花湖水库水温除22:00、2:00、6:00时0.5 m处水温略低于1—3 m处外,一昼夜间水温呈自上而下减小的趋势,一直有弱的分层现象,没有温跃层现象。白天8:00—18:00时0.5 m水温在25.4—27.5 ℃之间,平均26.5 ℃;1 m水温在25.1—26.4 ℃之间,平均25.7 ℃;2 m水温在24.0—26.0 ℃之间,平均25.0 ℃;3 m水温在23—25 ℃之间,平均24.5 ℃;4 m水温在21.5—24.0 ℃之间,平均23.0 ℃。夜间20:00—6:00时0.5 m水温在22.0—24.0 ℃之间,平均22.7 ℃;1 m水温在23.0—24.0 ℃之间,平均23.4 ℃;2 m水温在23—23.6 ℃之间,平均23.2 ℃;3 m水温在22.6—23 ℃之间,平均22.9 ℃;4 m水温在21.0—22.0 ℃之间,平均21.6 ℃。白天4 m及4 m以浅处水温随时间变化波动较大,夜间变化不明显。白天水柱温差较高,且波动较大,在4.5—8.5 ℃之间,夜间水柱温差较低,且波动较小,在4.0—6.2 ℃之间。

|
| 图 7 百花湖水库温度的时间深度等值图 Fig. 7 Depth-time distribution of water temperature in Baihua Reservoir |
Spearman相关分析表明(表 1),太阳入射光和水柱温差是影响浮游植物总丰度和湖泊假鱼腥藻丰度垂直分布格局昼夜变化的重要环境因素。水体中浮游植物总丰度的MI指数与太阳入射光有效辐射呈极显著正相关(r=0.794,n=10,P=0.01),与水柱温差呈显著正相关(r=0.759,n=10,P=0.05)。湖泊假鱼腥藻丰度的MI指数与太阳入射光有效辐射呈显著正相关(r=0.675,n=10,P=0.05),与水柱温差呈显著正相关(r=0.782,n=10,P=0.05)。
| 变量 Variation | 光合有效辐射/(μmol m-2 s-1) Photosynthetically available radiation | 水柱温差/℃ Water column temperature difference |
| *表示在0.05水平检验显著,**表示在0.01水平检验显著 | ||
| 浮游植物总丰度MI指数 Morisita′s index of phytoplankton densities | 0.794** | 0.759* |
| 湖泊假鱼腥藻丰度MI指数 Morisita′s index of Pseudanabaena limnetica densities | 0.675* | 0.782* |
具有伪空泡的蓝藻可以通过浮力调节机制改变自身在水体中的位置。在百花湖中,湖泊假鱼腥藻占据绝对优势地位,其垂直分布格局能够反映百花湖总体浮游植物的垂直分布情况。
3.1 浮游植物垂直分布格局研究期间百花湖浮游植物丰度白天大于夜间,与Olsson等关于藻类昼夜垂直迁移的研究结果一致[28]。湖泊假鱼腥藻为本研究的绝对优势种,白天具有明显的垂直迁移现象,已有研究证明具有伪空泡的蓝藻在水体中可以进行垂直运动[29],铜绿微囊藻(Microcystis aeruginosa) 、一种浮游蓝丝藻(Planktothrix rebescens)[30, 31]等具有伪空泡的水华蓝藻均存在垂直迁移现象。湖泊假鱼腥藻和浮游植物总丰度的昼夜垂直分布格局一致,在一昼夜间均呈聚集分布,且主要分布在0.5—4 m,白天聚集程度高于夜间。浮游蓝丝藻白天主要聚集在水体的2—3 m处[15],蓝藻在低光强下能够增加细胞气囊,使藻类群落占据湖泊的富光区域,为本研究中浮游植物主要聚集在4 m及4 m以浅水层进行了论证。
3.2 影响浮游植物垂直分布格局的因素分析Oliver R L,Walsby A E等对浮力变化的定量分析证实,蓝藻细胞浮力的调节涉及到影响细胞沉浮物(ballast)尤其是糖类的含量改变、细胞膨压升高引起伪空泡的破裂和伪空泡合成的调节3种机制[32, 33]。
碳水化合物为藻细胞内的主要镇重物[34],通过碳水化合物的积累与消耗影响细胞浮力的调节方式普遍存在于浮游蓝藻中,且能够迅速响应短时间内的环境变化。本研究中12:00时湖泊假鱼腥藻的总细胞丰度最大,而0.5 m处的细胞丰度却为除6:00外的最小值,为25.41×106个/L,最大值出现在2 m处,为62.31×106个/L。可能因为10:00—12:00光照充足,且光强逐渐增大,12:00时光强达到最大,湖泊假鱼腥藻通过光合作用积累的碳水化合物逐渐增多,这时糖类占细胞干重的比例可由30% 升至60%[3],细胞密度增大,由于伪空泡合成缓慢,因此伪空泡合成所提供的浮力不足以抵消糖类增加引起的细胞密度增大,细胞下沉,12:00之后光照逐渐减弱,积累的碳水化合物消耗到一定程度,细胞又开始上浮。Porat等提出了在浅水湖泊中糖含量变化是Aphanizomenon ovalisporum浮力变化主要原因的假设[35]。唐忠波等对浮游蓝丝藻的研究表明,藻丝的浮力对光照的响应可能通过藻细胞的糖含量变化实现[15]。
容易在浅水水体中形成优势的湖泊假鱼腥藻的伪空泡较脆弱,细胞膨压引起伪空泡破裂的浮力调节机制可能影响着湖泊假鱼腥藻的垂直迁移。临界压强是指导致伪空泡破裂的最小内外压强差。当伪空泡所受净压强高于临界压强时,会引起伪空泡破裂[36]。细胞内的伪空泡所受的净压强Pn=Ph+Pt+Pf-Pg。其中Ph为流体压强,Pt为细胞液与周围环境渗透势差引起的细胞内膨压,Pf为水柱上方气相压强,Pg为伪空泡内的气体压强。随着光照的增强,可溶性有机物积累增加,同时K+吸收增强,导致细胞膨压升高,伪空泡所受的净压强增大,当净压强大于临界压强时,伪空泡破裂,细胞浮力下降,细胞下沉。本研究中12:00时湖泊假鱼腥藻大量聚集在2m处,也可能与光照过强导致部分伪空泡破裂,细胞浮力下降有关。
本研究中8:00—18:00湖泊假鱼腥藻聚集程度及变化幅度较大,聚集程度最大的水层分别为3、0.5、2、0.5、1、0.5 m,8:00—18:00时0.5—2 m处光照强度的垂直变化程度较大,相同水层不同时间的光照强度差异明显,白天湖泊假鱼腥藻随光照强度的改变在水体中具有明显的垂直迁移现象。可见湖泊假鱼腥藻对光照变化的响应较快,而伪空泡合成过程缓慢,一般大于24h[37]。夜间假鱼腥藻的聚集程度及变化幅度不明显,与夜间无光相符。因此本研究中湖泊假鱼腥藻垂直迁移对光照的响应可能是通过碳水化合物含量变化和伪空泡破裂机制实现的。
在静水中,湖泊假鱼腥藻的垂直分布受浮力调节的控制,但水体扰动、风等因素也影响湖泊假鱼腥藻的分布。水柱上下层温差的变化对水库营养物质循环和水生态系统结构产生影响,从而形成利于蓝藻增殖和聚集的生境[38]。本研究中白天水柱温差较大,水体混合减弱,利于藻类在上层聚集,夜间温差减小,易导致水体混合,从而影响蓝藻的垂直分布,这也可能是导致湖泊假鱼腥藻的聚集程度白天高于夜间的原因之一。风是影响浮游植物垂向变化的关键因子之一,无风或弱风环境是研究藻类垂向迁移的基础,高风速可能是抑制蓝藻上浮、扩散的重要水文气象因素。本研究采样期间,天气晴朗无风,适合对藻类进行昼夜垂直分布特征进行研究。
| [1] | Sun R Y. Basic ecology. Beijing: Higher Education Press, 2002: 1-15. |
| [2] | Han B P, Lin X T, Li T. Study on current situation and Countermeasures of eutrophication in large and medium-sized reservoirs in Guangdong Province. Beijing: Science Press, 2003, 123-153. |
| [3] | Cheng H M, Qiu B S. Cyanobacterial Gas Vesicles and Their Regulation on the Vertical Distribution of Cyanobacteria in Water Body. Plant Physiology Communications, 2006, 42 (5): 974-980. |
| [4] | He Z H. Freshwater Ecology. Beijing: China Agriculture Press, 2000: 240-249. |
| [5] | Jones R I. Vertical distribution and diel migration of flagellates phytoplankton in a small humic lake. Hydrobiologia, 1988, 161(1): 75-87. |
| [6] | Yamamoto Y, Nakahara H. Seasonal variations in the diel vertical distribution of phytoplankton and zooplankton in a shallow pond. Phycological Research, 2006, 54(4): 280-293. |
| [7] | George D G, Heaney S I. Factors influencing the spatial distribution of phytoplankton in a small productive lake. Journal of Ecology, 1978, 66(1): 133-155. |
| [8] | Walsby A E,Reynolds C S,Oliver R L,Kromkamp J. The role of gas vacuoles and carbohydrate content in the buoyancy and vertical distribution of Anabaena minutissima in lake Rotongaio. Archiv für Hydrobiologie-Beiheft Ergebnisse der Limnologie,1989,32(1): 1-25. |
| [9] | Tadonleke R D, Jugnia L B, Simengando T, Zebaze S, Nola M. Short-term vertical distribution of phytoplankton populations in a shallow tropical lake (Lake Municipal, Yaoundé, Cameroon). Archiv für Hydrobiologie, 1998, 143(4): 469-485. |
| [10] | John D M, Whitton B A, Brook A J. The Freshwater Algal Flora of the British Isles: An Identification Guide to Freshwater and Terrestrial Algae. Cambridge: Cambridge University Press, 2002: 172-230. |
| [11] | Reynolds C S,Walsby A E. Water-blooms. Biological Reviews.,1975,50(4): 437-48l. |
| [12] | Ganf G G, Oliver R L. Vertical separation of light and available nutrients as a factor causing replacement of green algae by blue-green algae in the plankton of a stratified lake. Journal of Ecology, 1982, 70(3): 829-844. |
| [13] | Reynolds C S,Oliver R L,Walsby A E. Cyanobacterial dominance: the role of buoyancy regulation in dynamic lake environments. New Zealand Journal of Marine and Freshwater Research,1987,2 l(3): 379-390. |
| [14] | Spencer C N, King D L. Regulation of blue-green algal buoyancy and bloom formation by light, inorganic nitrogen, CO2, and trophic level interactions. Hydrobiologia, 1987, 144(2): 183-191. |
| [15] | Tang Z B, Chu Z S, Jin X C, Zeng Q R. Buoyancy Regulation and Vertical Distribution of Planktothrix mougeoti in the Simulator Experiment. Environmental science, 2008, 29(6): 1513-1517. |
| [16] | Thomas R H, Walsby A E. The efect of temperature on recovery of buoyancy by Microcystis. Microbilogy, 1986, 132(6):1665-1672. |
| [17] | Chen L L, Li Q H, Teng M D, Liu S P, Zhang M S. Cyan0bacteria composition and microcystins distribution of Wanfeng Reservoir and Baihua Reservoir on Guizhou Plateau. Ecology and Environmental Sciences, 2011, 20(6/7): 1068-1074. |
| [18] | Li Q H, Chen L L, Xia P H, Liu S P, Chen F F, Yu D M, Li C X. Structure of phytoplankton community and its relationship with environmental factors at the estuary of Maixi River in Baihua Reservoir, Guizhou Province. Journal Lake science, 2011, 23(4): 612-618. |
| [19] | Hu X H, Chen C, Li Y Y, Xiang G, Liu M S. An evaluation of water pollution and eutrophication in the Baihua Lake by means of phytoplankton. Journal of Guizhou Normal University: Acta Scientiarum Naturalium, 1999, 17(4): 1-7. |
| [20] | Hu H J, Wei Y X. The freshwarter algae of China. Beijing: Science Press, 2006: 1-948. |
| [21] | Wang Q X, Cao J G, Liu Y, Qin N. Altas of Algae in Shanghai Jiuduansha Wetland Nature Reserve and Its Adjacent Waters. Beijing: Science Press, 2008: 1-160. |
| [22] | Zhang Z S,Huang X F. Method for the study of freshwater plankton. Beijing: Science Press,1995: 1-414. |
| [23] | Thackeray S J, George D G, Jones R I, Winfield I J. Statistical quantification of the effect of thermal stratification on patterns of dispersion in a freshwater zooplankton community. Aquatic Ecology, 2006, 40: 23-32. |
| [24] | Aguilar A. Adult survival and movement in males of the damselfly Hetaerina Cruentata (Odonata:Calopterygidae). Florida Entomologist, 1994, 77(2): 256-264. |
| [25] | Hills J M, Thomason J C. A multi-scale analysis of settlement density and pattern dynamics of the barnacle Semibalanus balanoides. Marine Ecology Progress Series, 1996, 138(1/3): 103-115. |
| [26] | Quinn V S, Graves B M. Space use in Response to conspecifics by the red-backed salamander(Plethodon einereus,Plethodontidae,Caudata). Ethology, 1999, 105(11): 993-1002. |
| [27] | Ye L. Studies on the Eutrophication and the Spring Phytoplankton Bloom in Xiangxi Bay of Three-Gorge Reservoir[D]. Wuhan: Chinese Academy of Sciences,2006. |
| [28] | Olsson P, Graneli E. Observations on diurnal vertical migration and phased cell division for three coexisting marine dinoflagellates. Jouranl of Plankton Research, 1991, 13(6): 1313-1324. |
| [29] | Li X P. Cyanobacteria Movements and Its Bloom Control. Chinese Journal of Nature,2008,30(5): 280-286. |
| [30] | Bright D I, Walsby A E. The daily integral of growth by Planktothrix rubescens calculated from growth rate in culture and irradiance in Lake Zurich. New Phytologist, 2000, 146(2): 301-316. |
| [31] | Wallace B B, Bailey M C, Hamilton D P. Simulation of vertical position of buoyancy regulating Microcystis aeruginosa in a shallow eutrophic lake. Aquatic Science, 2000, 62(4): 320-333. |
| [32] | Oliver R L. Floating and sinking in gas-vacuolate cyanobacteria. Journal of Phycology, 1994, 30(2): 161-173. |
| [33] | Walsby A E. Gas vesicles. Microbiological Reviews, 1994:94-l44. |
| [34] | Zhang H C,Chen X C,Li C J. The effect of light intensity on cyanophyta vertical distribution. Environmental pollution and control,2010,32(5):64-67. |
| [35] | Porat R, Teltsch B, Perelman A, Dubinsky Z. Diel buoyancy changes by the cyanobacterium Aphanizomenon ovalisporum from a shallow reservoir. Journal of P1ankton Research, 2001, 23(7): 753-763. |
| [36] | Walsby A E. The elastic compressibility of gas vesicles. Proceeding the Royal of Society London Biological, 1982, 216(1204): 355-368. |
| [37] | Hayes P K, Walsby A E. An investigation into the recycling of gas vesicle protein derived from collapsed gas vesicles. Microbio1ogy, 1984, 130(6): 1591-l596. |
| [38] | Gao T H,Qian X,Chu Z S,Wang S,Wu J,Chuai X M,Yang L Y. Effect of Climate,Hydrological and Ecological Processes on Yanghe Reservoir′s Eutrophicaiton. Journal of Hydroecology,2010,31(3): 28-31. |
| [1] | 孙儒永. 基础生态学. 北京: 高等教育出版社, 2002: 1-15. |
| [2] | 韩博平, 林旭钿, 李铁, 广东省大中型水库富营养化现状与防治对策研究. 北京: 科学出版社, 2003, 123-153. |
| [3] | 成慧敏, 邱保胜. 蓝藻的伪空泡及其对蓝藻在水体中垂直分布的调节. 植物生理学通讯, 2006, 42 (5): 974-980. |
| [4] | 何志辉. 淡水生态学. 北京:中国农业出版社, 2000: 240-249. |
| [15] | 唐忠波, 储昭升, 金相灿, 曾清如. 孟氏浮游蓝丝藻在模拟水体中的垂直分布与浮力规律研究. 环境科学, 2008, 29(6): 1513-1517. |
| [17] | 陈丽丽, 李秋华, 滕明德, 刘送平, 张明时. 两座高原水库蓝藻群落结构与微囊藻毒素的分布对比研究. 生态环境学报, 2011, 20(6/7): 1068-1074. |
| [18] | 李秋华, 陈丽丽, 夏品华, 刘送平, 陈峰峰, 余德民, 李存雄. 贵州百花湖麦西河河口浮游植物群落结构及与环境因子关系. 湖泊科学, 2011, 23(4): 612-618. |
| [19] | 胡晓红, 陈椽, 李银燕, 向刚, 刘美珊. 以浮游植物评价百花湖水质污染及富营养化. 贵州师范大学学报: 自然科学版, 1999, 17(4): 1-7. |
| [20] | 胡鸿均, 魏印心. 中国淡水藻类. 北京: 科学出版社, 2006: 1-948. |
| [21] | 王全喜, 曹建国, 刘妍, 钦娜. 上海九段沙湿地自然保护区及其附近水域藻类图集. 北京: 科学出版社, 2008: 1-160. |
| [22] | 章宗涉, 黄祥飞. 淡水浮游生物研究方法. 北京: 科学出版社, 1995: 1-414. |
| [27] | 叶鳞. 三峡水库香溪河库湾富营养化及春季水华研究[D]. 武汉: 中国科学院, 2006. |
| [29] | 李小平.蓝藻运动与水华早期预防和控制.自然杂志, 2008, 30(5): 280-286. |
| [34] | 张海春, 陈雪初, 李春杰. 光照度对蓝藻垂直迁移特性影响研究. 环境污染与防治, 2010, 32(5): 64-67. |
| [38] | 高天珩, 钱新, 储昭升, 王晟, 武俊, 揣小明, 杨柳燕. 气候、水文和生态过程对洋河水库富营养化影响研究. 水生态学杂志, 2010, 31(3): 28-31. |