 2014, Vol. 34
2014, Vol. 34文章信息
- 李伟, 张翠萍, 魏润鹏
- LI Wei, ZHANG Cuiping, WEI Runpeng
- 广东中西部桉树人工林植物多样性与林龄和土壤因子的关系
- Relationship of under-story vegetation diversity with stand age and soil factor of eucalypt plantations in central-western Guangdong
- 生态学报, 2014, 34(17): 4957-4965
- Acta Ecologica Sinica, 2014, 34(17): 4957-4965
- http://dx.doi.org/10.5846/stxb201301060041
-
文章历史
- 收稿日期:2013-1-9
- 网络出版日期: 2014-3-5






2. 华南农业大学林学院, 广州 510642;
3. 中国林业科学研究院林业新技术所, 北京 100091;
4. 慧霸达生资有限公司, 中国香港 999077
2. Forestry College, South China Agricultural University, Guangzhou 510642, China;
3. Research Institute of Forestry New Technology, Chinese Academy of Forestry, Beijing 100091, China;
4. Weirptech Bioresources Limited, Hong Kong 999077, China
我国高投入桉树人工林于20年前开始出现,近十年来快速发展。随着栽培品种、苗木繁殖、栽培技术、经营目的、采伐更新等内容逐步趋向一致,桉树人工林在我国南方已成独具一格的森林生态系统。然而,其快速发展的同时也带来一些生态、环境、社会等方面的潜在问题,并日益受到人们关注[1, 2, 3],林下植物多样性便是主要关注点之一。相关研究表明桉树种植导致林下植物多样性的减少[4, 5],但也有调查持不同的结论,指出有利本土植被和物种的恢复[6, 7]。综合相关研究可以发现,桉树林下植物多样性的变化是受自然环境因素(如原植物群落类型、立地条件)和人为因素(包括立地条件不适、栽培和经营管理措施不当、取走枯枝落叶等)等因素造成的[4, 7, 8]。桉树人工林分布广阔,现有林下植物多样性调查研究通常局限于特定环境条件,在广东的中西部低丘地区,依然缺少相关的研究报道。
有关研究发现在林分不同年龄阶段,林下植被的物种组成和数量都有很大的变化,而且不同林分林下植被的物种多样性随林木发育阶段出现不同的变化规律[9, 10]。目前国内桉树人工林多为短轮伐期(5—7a),关于林下植被随年龄变化的研究报道很少[11],而国外的报道主要集中10a以上的林分[12, 13]。土壤是植物生长发育的重要物质基础,其理化性质与植物多样性有密切的关系[14]。由于土壤对多样性的影响较复杂,研究结果也缺乏一致的规律性,而现有的研究多涉及森林群落[14]、草原群落[15]以及盐沼沙生植被[16],对桉树人工林土壤因子与林下植物多样性关系的研究却没有相关报道。
本文以桉树人工林为研究对象,通过样地调查,分析不同龄级林分林下物种多样性的变化规律;运用CCA排序方法综合分析植被分布与土壤因子关系,探讨植物多样性与环境因素之间关系,从而研究规模种植桉树对当地植被影响的作用机制。
1 研究地区概况和方法 1.1 研究地区概况高要市位于东经112°26′—112°52′、北纬22°47′—23°26′,地处广东省中部偏西、西江中下游。地貌走势西北高、东南低,境内山岗、河谷、丘陵、平原交错,西北部属山区和半山区,南部属丘陵区,而大多数桉树人工林林地低于海拔400 m。主要成土母岩为花岗岩、砂岩等酸性基岩,有机质相对贫瘠。属亚热带季风气候,根据市气象局资料,年平均气温22 °C,年平均积温8041 °C,无霜期340 d;年平均降雨量1647.9 mm,4—9月降雨量最多,占全年降雨量83%,历年平均降雨日数156 d;历年平均日照时数1801.6 h,年平均湿度80%左右。
1.2 研究方法 1.2.1 样地设置和调查方法按林龄1—2、3—4和5—6年生分3个龄级,每个龄级分别选择独立分布的林分,每林分于上、中和下坡以及谷沟处各布设有代表性样地1个,样地规格15 m×15 m,上下边界沿等高线走向,共计72个样地,用于林下乔木层植被调查。在每个入选样地内,以样地中心位置,沿坡向方向,靠样地上下边界设置边长为4 m的方形样方各1个,用于灌木层植被调查;在每个样地内,从中心点起,沿水平及垂直方向,分别向左、右、上、下,靠近样地边界设置边长为1 m的方形小样方各1个,用于草本层植被调查。记录植物物种名称、个数、高度、盖度等指标。因为抚育铲劈等原因,绝大多数乔木树种处于林下灌丛或低层状态,因此把乔木树种归入灌木层分析。
所有入选林分(样地)造林措施基本一致,包括造林前炼山、清山、整地、种植穴规格、无性苗木等,林木株行距均为2 m×3 m;第1年造林前每株施相同的专用复合基肥375 g,第1和2年各除草1次,第2和3年每株各施相同专用复合追肥500 g;林木生长状况良好,保存率达90%以上,第2年后林分树冠基本趋向郁闭。
1.2.2 土壤取样及测定方法在每龄级24个样地中,进一步选取6个典型样地,共计18个样地,进行土壤取样和相关因子测定。在每样地中心和四角处5点各挖60 cm深的土壤剖面,按照不同深度(0—20 cm、20—40cm、40—60cm)分层取样,5位置相应土层土样混合均匀后带回实验室。风干土样过2 mm筛子,研磨后用100目分子筛再过筛。土样测定项目和方法如下:环刀法测定土壤容重、电位法测定土壤pH值、重铬酸钾氧化外加热法测定有机质含量、凯氏定氮法测定土壤全氮含量、钼锑抗比色法测定全磷和NaOH熔融法测定全钾含量。
1.3 数据处理和统计分析 1.3.1 物种累计曲线物种累积曲线(species accumulation curve)用于描述随着抽样量的加大物种增加的状况,是理解调查样地物种组成和预测物种丰富度的有效工具,在生物多样性和群落调查中,被广泛用于抽样量充分性的判断以及物种丰富度估计[17]。采用一阶折刀指数获得期望物种丰富度(Sp),计算公式如下[18]:
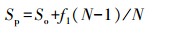
对每块样地,分别计算每种灌木、草本或藤本物种的重要值,计算公式为:重要值(%) =(相对盖度+相对频度+相对密度)/3。
1.3.3 物种多样性物种丰富度S=出现在样地内的物种数
Shannon指数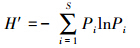
Simpson指数 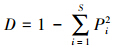
Pielou′s指数 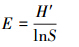
式中,Pi为第i个物种种群占总群体的比值,∑Pi=1。
以18个典型样地植物重要值和土壤测定数据为基础,首先构建72×18维物种重要值矩阵和6×18维土壤因子矩阵,利用CANOCO 4.5软件,进行典范对应分析(CCA)[20];然后采用SAS软件中CORR模块计算4个多样性指数与土壤因子的简单相关系数。因为采用样土不同分层与其均值所得出的结果类似,在后面有关分析中仅涉及不同土壤分层均值的结果。
1.3.5 方差分析分析3个龄级之间物种多样性指数的差别,采用SAS程序包中的单因素等重复试验模型进行方差分析。
2 结果与分析 2.1 桉树林下植物种类组成本次研究调查的72样地中,共有维管束植物136种,隶属于54科107属,其中灌木植物80种,占所调查物种总数的58.8%,隶属于37科68属;草本层植物36种(26.5%),分属15科32属;藤本有20种,占物种总数的14.7%,分属13个科17个属。说明桉树林下植被中灌木层所占种类最多,草本层次之,藤本最少(表 1)。
| 植物类型 Plant type | 科 Family | 科占比例/% Family′s proportion | 属Genus | 属占比例/% Genus′ proportion | 种 Species | 种占比例/% Species′ proportion |
| 灌木Shrub | 37 | 56.9 | 68 | 58.1 | 80 | 58.8 |
| 草本Herbaceous | 15 | 23.1 | 32 | 27.4 | 36 | 26.5 |
| 藤本Liana | 13 | 20.0 | 17 | 14.5 | 20 | 14.7 |
表 2中列出了各植物类型中重要值排名前5位的灌木、草本和藤本。从表中可以看出,灌木层主要组成种包括桃金娘(Rhodomyrtus tomentosa (Ait.) Hassk.)、鬼灯笼(Clerodendrum fortunatum L.)、山黄麻(Trema tomentosa (Roxb.) Hara)、山苍子(Litsea cubeba (Lour.) Pers.)、野牡丹(Melastoma candidum D. Don)等,这些物种出现频率较高,重要值都在44%以上,其余灌木种在样地里出现频率不高,零星的出现于各个样地或群生在单一样地中。草本层中芒萁(Dicranopteris dichotoma (Thunb.) Bernh.)的重要值最高,为优势物种,其余主要物种包括芒草(Miscanthus sinensis Anderss.)、乌毛蕨(Blechnum orientale L.)、海金沙(Lygodium japonicum (Thunb.) Sw.)等,重要值在53%以上。藤本物种中酸藤子(Embelia laeta (L.) Mez)、玉叶金花(Mussaenda pubescens Ait. f.)、菝葜(Smilax china L.)和无根藤(Cassytha filiformis Linn.)出现的频率较高,为主要物种,重要值在4%以上。在所有植物种中,草本中的芒萁和芒草的重要值排在前两位,分别为252%和112%,说明芒萁和芒草是桉树林下植物的优势种(表 2)。
| 植物类型 Plant type | 种名 Species name | 重要值/% Important value |
| 灌木Shrub | 桃金娘Rhodomyrtus tomentosa | 70.4 |
| 鬼灯笼Clerodendrum fortunatum | 61.3 | |
| 山黄麻Trema tomentosa | 54.9 | |
| 山苍子Litsea cubeba | 49.1 | |
| 野牡丹Melastoma candidum | 44.3 | |
| 草本 | 芒萁Dicranopteris dichotoma | 251.8 |
| Herbaceous | 芒草Miscanthus sinensis | 111.8 |
| 乌毛蕨Blechnum orientale | 70.9 | |
| 海金沙Lygodium japonicum | 61.2 | |
| 小叶海金沙Lygodium scandens (Linn.) Sw. | 53.4 | |
| 藤本Vine | 酸藤子Embelia laeta | 45.8 |
| 玉叶金花Mussaenda pubescens | 32.6 | |
| 菝葜Smilax china | 16.0 | |
| 无根藤Cassytha filiformis | 9.0 | |
| 锡叶藤Tetracer aasiatica (Lour.) Hoogl. | 4.2 |
3个龄级桉树林下植物种数随样地数累积的结果如图 1。随着样地数增加,种数累积曲线在急剧上升后变化趋缓,表明抽样充分,可进行数据分析[17]。图 1表明随着样地数增加,种数也不断增加,在前8个样地中,3个龄级林下植物种数不存在差异,但样地数超过9个以后,1—2年生龄级林下植物种数在一定程度上稳定地少于另两个龄级,而3—4年生龄级稍低于5—6年生龄级,但是两者没有显著差异。

|
| 图 1 基于样本的物种累积曲线 Fig. 1 Sample-based species accumulation curves |
方差分析表明,各种多样性指数在各个龄级之间差异统计上不显著(P>0.05)。但是,3个龄级之间仍呈现一定的变化趋势(图 2),1—2年生桉树林下植物多样性均低于其它两个龄级,Shannon、Simpson和丰富度指数在3—4年生时最高,虽然3—4年和5—6年生两个龄级林分之间没有统计上的差异。这种差别和变化趋势可能是短轮伐期林龄(6a)影响的结果,如果轮伐期长,会变成更加明显[13],因为人为抚育干扰的减少,林下物种结构也会趋向稳定。

|
| 图 2 不同龄级下物种多样性指数 Fig. 2 Diversity index of different ages |
对桉树林下植被物种重要值与土壤中全氮、全磷、全钾、pH值、容重和有机质进行CCA排序分析。结果表明(图 3和表 3),CCA第一和第二排序轴与土壤中有机质、全磷、全钾和容重等因子显著相关,特征值为0.244和0.181,物种与前2个排序轴的相关系数分别为0.856和0.943,两者的方差累计贡献率达到51.1%。在6个环境因子中,对第一排序轴影响最大的是全磷,分量系数为-0.6301,其次是有机质,分量系数为-0.5315,均达极显著水平,而pH值、全钾、全氮和容重的作用较小,没有达到明显程度(表 3)。对第二轴影响极显著的因子有全钾和土壤容重,分量系数分别为0.5841和-0.5425,pH值的作用较大但没达到显著水平,分量系数为0.4418,其它因子作用比较小。

|
| 图 3 群落优势种CCA排序图 Fig. 3 CCA ordination diagram of dominant species of forest community 图中仅显示权重5%—100%的物种, 1: 桃金娘; 3: 乌桕; 4: 鬼灯笼; 11: 山黄麻; 12: 了哥王; 14: 山苍子; 16: 春花; 17: 野牡丹; 21: 豺皮樟; 22: 紫玉盘(Uvaria macrophylla Roxb.); 27: 三叉苦; 29: 假鹰爪(Desmos chinensis Lour.); 28: 无根藤; 30: 厚皮香(Ternstroemia gymnanthera (Wight et Arn.) Bedd.); 31: 黄牛木; 32: 芒萁; 33: 芒草; 34: 黄药; 38: 酸藤子; 39: 白背叶; 42: 海金沙; 48: 梅叶冬青; 49: 毛相思子; 58: 黄毛楤木; 62: 玉叶金花; 68: 假地豆 |
| 土壤因子 Soil factor | 第一排序轴 Axis1 | 第二排序轴Axis2 | 第三排序轴 Axis3 | 第四排序轴 Axis4 |
| *P< 0.05,**P< 0.01,***P< 0.001 | ||||
| pH值 pH value | 0.2015 | 0.4418 | -0.1493 | -0.1513 |
| 有机质 Organic matter | -0.5315* * | -0.2260 | -0.2476 | -0.6965* * * |
| 全钾 Total potassium | 0.3497 | 0.5841* * * | -0.3907 | -0.0446 |
| 全磷 Total phosphorus | -0.6301* * * | 0.3154 | -0.4210 | -0.3149 |
| 全氮 Total nitrogen | -0.3270 | -0.1527 | -0.3097 | 0.0880 |
| 容重 Bulk density | -0.3278 | -0.5425* * | 0.1357 | 0.6938* * * |
| CCA排序概要 Summary of CCA ordination | ||||
| 特征值 Eigenvalue | 0.244 | 0.181 | 0.146 | 0.135 |
| 物种与土壤相关 Species and soil correlation | 0.856 | 0.943 | 0.898 | 0.949 |
| 物种与土壤相关的方差累积比例 Cumulative percentage variance of species-soil correlation /% | 29.4 | 51.1 | 68.7 | 84.9 |
| 所有典范轴的显著性检验 Test of significance of all canonical axes | P=0.037 | |||
沿第一排序轴由左至右,全钾、全氮和pH值逐渐增加,有机质、全磷和容重则呈相反趋势,逐渐减少。有机质和全氮沿第二轴由下到上逐渐减少,pH值、全磷、全钾和容重则由下到上逐渐增大。排序图(图 3)上位于第一轴右上端的物种,如毛相思子(Abrus mollis Hance)分布主要受到土壤中pH值的影响,在pH值相对较大的区域分布较多;了哥王(Wikstroemia indica (L.) C. A. Mey)和豺皮樟(Litsea rotundifolia Hemsl. var. oblongifolia (Nees) Allen)分布主要受到全氮的影响,在全氮含量较高的区域分布较多;白背叶(Mallotus apelta (Lour.) Muell. Arg.)、梅叶冬青(Ilex asprella (Hook. et Arn.) Champ. ex Benth.)、黄毛楤木(Aralia decaisneana Hance)分布在第一轴的左上端,即在土壤容重和全 磷含量高的环境条件下分布较多;黄药(Rhamnus crenata Sieb. et Zucc.)、假地豆(Desmodium heterocarpon (Linn.) DC.)、春花(Raphiolepis indaca L.)、乌桕(Sapium sebiferum (L.) Roxb.)等受有机质和全氮含量影响较大,而桃金娘、鬼灯笼、山黄麻、山苍子、野牡丹、三叉苦、黄牛木(Cratoxylum cochinchinense (Lour.) Bl.)、芒萁、芒草、酸藤子、玉叶金花等物种广泛分布于各个样地内,受土壤因子影响较小,适应范围较宽,因此位于排序图中心位置。
2.5 物种多样性指数与土壤因子的关系从表 4中可见,丰富度指数与全氮之间的相关达到显著性水平,Simpson指数与全氮以及Pielou′s指数与pH值之间接近显著水平(P=0.05,r0=0.468),Simpson、Shannon和丰富度指数与pH值之间存在次级显著关系(P=0.1,r0=0.400),Shannon和Pielou′s指数与全氮之间的相关系数也较大,所有这些关系均为正相关(表 4),说明在6个土壤因子中全氮和pH值对林下植被物种多样性的影响比较大,也最为稳定,其它4个土壤因子的作用偏低。
| 多样性指数 Diversity Index | pH值 | 有机质Organic matter | 全钾 Total Potassium | 全磷 Total phosphorus | 全氮 Total nitrogen | 容重 bulk density |
| *P < 0.05 | ||||||
| Simpson指数Simpson index | 0.401 | -0.262 | 0.141 | 0.149 | 0.462 | 0.233 |
| Shannon指数 Shannon index | 0.418 | -0.211 | 0.102 | 0.102 | 0.343 | 0.229 |
| 丰富度指数 Richness | 0.401 | -0.219 | 0.128 | 0.128 | 0.559* | 0.263 |
| Pielou′s指数Pielou′s index | 0.466 | -0.258 | 0.052 | 0.052 | 0.367 | 0.273 |
本研究所调查的72个桉树林分样地中,林下植物种数136种,隶属于54科107属,植被结构主要有两种类型,即灌木-草本类型和纯草本型,植物种数目以灌木为主,草本层次之,但是芒萁和芒草为优势种。商业桉树人工林是过去20年来逐步兴起的,林下植被群落受标准化集约经营影响较大[21, 22],但是植被物种组成数量上的差别仍可能与地区水热条件、原植被群落、土壤条件、树种年龄等因素有关[7, 13]。在桉树林下植物种数组成上,广东中西部比海南岛(224种)明显偏低[23],比雷州半岛(72种)显著丰富[21],与云南省西南部(135种)接近[7]。
桉树林下植物数目有随着年龄的提高而增加的趋势,而多样性指数则在3—4年生时最高,这一现象和一些研究结论类似[5, 11, 13],但是与个别报道不同[21],这一结果可能反映了广东中西部短轮伐期桉树人工林的实际情况。虽然造林前林地经过炼山、清山和整地,但林地仍存在不少植物的繁殖体,如被埋在土壤中的根蔸和种子库[24],这些植物尤其是原来群落中的优势种具有极强的无性或有性繁殖能力。当环境条件有利于其生长时,会很快恢复生长,重新占据林地空间。所以在桉树造林后不到3a,林下植物种类显著增加或者快速恢复[11, 25],而到林分后期(5—6年生),林木直径和树高生长大大减慢,多数林分的林冠反而变成稀疏,有利林下植物的恢复和重新定居。
CCA排序是将植物种数据矩阵与环境因子矩阵相结合,能较好地表达群落的环境梯度,有利于对排序轴的生态意义进行解释[20]。在本研究的CCA第一排序轴中,土壤有机质和全磷含量的影响极为显著,两者均具负相关的作用,即有机质和全磷含量越高,排序轴值越低(表 3)。在第二排序轴中,土壤中全钾与容重的作用极为显著,前者正相关,后者负相关,pH值具有一定程度但不显著的正相关作用(表 3)。不同植物对土壤条件需求存在差异,有自己特定的分布中心和分布范围(图 3),这可以反映出群落类型的分布。
由于调查林分的地理分布变化不大,样地的气候环境相似,所以由地理位置变化所引起的热量、水分等气候因素差异对多样性的影响不是很大,本文主要分析土壤因素所引起的多样性指数的变化。在所调查的土壤因子中,pH值和全氮与各种植物多样性指数之间均具有比较密切的正向关系,其中,只有全氮与与丰富度指数之间达到统计上的显著性水平(P<0.05),全氮与Simpson指数和pH值与Pielou′s指数之间的关系接近显著水平(表 4)。可以认为,土壤pH值和全氮含量对高要市桉树林下植被物种多样性影响较大,土壤肥沃和酸碱度适中有利于短周期人工林林下植被物种多样性的恢复和维持。
大面积连片栽培的人工林对林下生物多样性存在一定的负面影响,这是人工林经营中的一个普遍现象[4, 5, 23]。目前国内大多数桉树林分的轮伐期为6a上下,属于高度集约经营的森林,一般情况下,在前两年实行比较强化的营林措施,包括修路、清山、炼山、整地、除草施肥,第3年施肥不除草,以后不再施肥抚育,最后修路、采伐和运输。在连续经营中,对林地的人为干扰变得十分频繁,这些干扰对林下植被群落的保护基本上是不利的,物种多样性会随之下降[13, 26]。由于时间短,在单一轮伐期内,林下植被难于平稳地恢复、演进并形成稳定的植物群落,因此,现有的桉树林下植物多样性随林龄的变化可能局限于特定的时空背景[11, 13]。另一方面,在不少水土流失和植被破坏严重地区,由于适应性强,桉树种植对当地植物多样性修复或恢复又有相当的正面意义[8, 27],至少在一定的时限内如此。
综合不同调查研究结果来看,桉树林下植物物种多样性的变化或减少,是自然环境(如原植物群落类型和立地条件)和人为因素(栽培和经营管理措施不当、取走枯枝落叶等)综合影响的结果,只是在不同的地区,各个因素作用的方式和程度不同[7, 13, 25, 28]。桉树栽培主要是通过改变环境条件和资源的可利用性,如土壤水分、养分以及营养结构而对本地物种产生间接的影响,减少了物种多样性。
致谢: 沐海涛、张启明、李仁军等协助野外植被调查和土壤取样,许建新、曹鹤等在植物识别和土壤实验方面提供帮助,特此致谢。| [1] | Shiva V, Bandyopadhyay J. Eucalyptus -a disastrous tree for India. Ecologist, 1983, 13(5): 184-187. |
| [2] | Wei R P. Genetic diversity and sustainable productivity of eucalypt plantations in China//Changing Patterns-Tree Introduction and Phytogeography. Beijing: China Forestry Publishing House, 2005: 19-27. |
| [3] | Xu D P, Zhang N N. Study development of ecological effect of Eucalyptus plantation. Guangxi Forestry Science, 2007, 35(4): 179-187, 201-201. |
| [4] | Han Y S, Wei Y C, Ouyang Z Y, Cao Y. Effects of continuous planting rotation on forest structural characteristics and water holding capacity of Eucalyptus plantations. Acta Ecologica Sinica, 2008, 28(9): 4609-4617. |
| [5] | Wen Y G, Liu S R, Chen F. Effects of continuous cropping on understorey species diversity in eucalypt plantations. Chinese Journal of Applied Ecology, 2005, 16(9): 1667-1671. |
| [6] | Pan H, Huang S D, Zhang J W, Wang G H, Zhu H R. Ecological defects and countermeasures of eucalyptus plantation in Fujian Province. Chinese Journal of Eco-Agriculture, 2009, 17(3): 605-609. |
| [7] | Liu P, Qin J, Liu J C, Wang H F, Wang X K. Comparison of structure and species diversity of Eucalyptus community. Acta Ecologica Sinica, 2011, 31(8): 2227-2235. |
| [8] | Alem S, Pavlis J. Native woody plants diversity and density under Eucalyptus camaldulensis plantation, in Gibie Valley, South Western Ethiopia. Open Journal of Forestry, 2012, 2(4): 232-239. |
| [9] | Gao R H, Dong Z, Zhang H, Li J Q. Study on regeneration process and biodiversity characteristic of Populus euphratica community in the Ejina Natural Reserve, Inner Mongolia of China. Acta Ecologica Sinica, 2005, 25(5): 1019-1025. |
| [10] | Zhao X H, Zhang C Y, Zheng J M. Correlation between gap structure and tree diversity of mixed-broad-leaved Korean pine forests in northeast China. Chinese Journal of Applied Ecology, 2005, 16(12): 2236-2240. |
| [11] | Gu L H, Zhou Y. Vegetation structure and species diversity in eucalypt understory at different stand age. Guangdong Forestry Science and Technology, 2012, 28(1): 46-52. |
| [12] | Tyynelä T M. Species diversity in Eucalyptus camaldulensis woodlots and miombo woodland in Northeastern Zimbabwe. New Forests, 2001, 223(3): 239-257. |
| [13] | Carneiro M, Fabião A, Martins M C, Fabião A, Abrantes da Silva M, Hilário L, Lousã M, Madeira M. Effects of harrowing and fertilisation on understory vegetation and timber production of a Eucalyptus globulus Labill. plantation in Central Portugal. Forest Ecology and Management, 2008, 255(3/4): 591-597. |
| [14] | Yang W Q, Zhong Z C, Tao J P. The distribution of soil available phosphorus and its correlations with plant species diversity in the forest of Mt. Jinyun. Chinese Journal of Ecology, 2001, 20(4): 24-27. |
| [15] | Bai Y F, Li L H, Wang Q B, Zhang L X, Zhang Y, Chen Z Z. Changes in plant species diversity and productivity along gradients of precipitation and elevation in the Xilin River basin, Inner Mongolia. Acta Phytoecol Sinica, 2000, 24(6): 667-673. |
| [16] | Thalen D C P. Variation in some saltmarsh and dune vegetations in the Netherlands with special reference to gradient situations. Acta Botanica Neerlandica, 1971, 20: 321-342. |
| [17] | Li Q, Chen Y Q, Xu Z H. Research methods on ant community. Chinese Journal of Ecology, 2009, 28(9): 1862-1870. |
| [18] | Heltshe J F, Forrester N E. Estimating species richness using the jackknife procedure. Biometrics, 1983, 39(1): 1-11. |
| [19] | Colwell R K. EstimateS: Statistical estimation of species richness and shared species from samples. Version 8.2, http://pur1.ocle.org/estimates, 2009. |
| [20] | Zang R G, Jing X H, Ding Y, Cheng K W, Bai Z Q, Zhang X P, Guo Z J, Zhang W Y. Quantitative classification, ordination and environmental analysis of woody plant communities in Xiaodonggou forest area of the Altai Mountain, Xinjiang. Scientia Silvae Sinicae, 2010, 46(2): 24-31. |
| [21] | Yu X B, Zhong L S, Yang W D, Long T. Structure of understorey vegetation in Eucalyptus plantations. Chinese Journal of Tropical Crops, 1999, 20(1): 66-72. |
| [22] | Nagai M, Yoshida T. Variation in understory structure and plant species diversity influenced by silvicultural treatments among 21-to 26-years old Picea glehnii plantations. Journal of Forest Research, 2006, 11(1): 1-10. |
| [23] | Yang Z H. A Comparative Study on Understorey Vegetation Diversity of Eucalyptus Plantation in Hainan Island . Haikou: South China University of Tropical Agriculture, 2001. |
| [24] | Egler F E. Vegetation science concepts I. initial floristic composition, a factor in old-field vegetation development with 2 figs. Vegetatio, 1954, 4(6): 412-417. |
| [25] | He Y L, Fu H Y. Review of studies on understorey of plantations. Forest Research, 2002, 15(6): 727-733. |
| [26] | Peacock R J. Vegetation Biodiversity Response of Eucalyptus Regrowth Forest to Thinning and Grazing. Sydney: Macquarie University, 2008. |
| [27] | Yirdaw E, Luukkanen O. Indigenous woody species diversity in Eucalyptus globulus Labill. ssp. globulus plantations in the Ethiopian highlands. Biodiversity and Conservation, 2003, 12(3): 567-582. |
| [28] | Burrows N D, Ward B, Cranfield R. Short-term impacts of logging on understorey vegetation in a jarrah forest. Australian Forestry, 2002, 65(1): 47-58. |
| [2] | 魏润鹏. 中国桉树人工林的遗传多样性与长期生产力的可持续性 // 格局在变化-树木引种与植物地理. 北京: 中国林业出版社, 2005: 19-27. |
| [3] | 徐大平, 张宁南. 桉树人工林生态效应研究进展. 广西林业科学, 2007, 35(4): 179-187, 201-201. |
| [4] | 韩艺师, 魏彦昌, 欧阳志云, 曹云. 连栽措施对桉树人工林结构及持水性能的影响. 生态学报, 2008, 28(9): 4609-4617. |
| [5] | 温远光, 刘世荣, 陈放. 连栽对桉树人工林下物种多样性的影响. 应用生态学报, 2005, 16(9): 1667-1671. |
| [6] | 潘辉, 黄石德, 张金文, 王光华, 朱洪如. 试论福建省桉树人工林的生态问题及其对策. 中国生态农业学报, 2009, 17(3): 605-609. |
| [7] | 刘平, 秦晶, 刘建昌, 王华锋, 王效科. 桉树人工林物种多样性变化特征. 生态学报, 2011, 31(8): 2227-2235. |
| [9] | 高润宏, 董智, 张昊, 李俊清. 额济纳绿洲胡杨林更新及群落生物多样性动态. 生态学报, 2005, 25(5): 1019-1025. |
| [10] | 赵秀海, 张春雨, 郑景明. 阔叶红松林林隙结构与树种多样性关系研究. 应用生态学报, 2005, 16(12): 2236-2240. |
| [11] | 古丽红, 周毅. 不同林龄桉树林下植被结构与物种多样性. 广东林业科技, 2012, 28(1): 46-52. |
| [14] | 杨万勤, 钟章成, 陶建平. 缙云山森林土壤速效P的分布特征及其与物种多样性的关系研究. 生态学杂志, 2001, 20(4): 24-27. |
| [15] | 白永飞, 李凌浩, 王其兵, 张丽霞, 张焱, 陈佐忠. 锡林河流域草原群落植物多样性和初级生产力沿水热梯度变化的样带研究. 植物生态学报, 2000, 24(6): 667-673. |
| [17] | 李巧, 陈又清, 徐正会. 蚂蚁群落研究方法. 生态学杂志, 2009, 28(9): 1862-1870. |
| [20] | 臧润国, 井学辉, 丁易, 成克武, 白志强, 张新平, 郭仲军, 张炜银. 新疆阿尔泰山小东沟林区木本植物群落数量分类, 排序及其环境解释. 林业科学, 2010, 46(2): 24-31. |
| [21] | 余雪标, 钟罗生, 杨为东, 龙腾. 桉树人工林林下植被结构的研究. 热带作物学报, 1999, 20(1): 66-72. |
| [23] | 杨再鸿. 海南岛桉树人工林林下植物多样性的比较研究 [D]. 海口: 华南热带农业大学, 2001. |
| [25] | 何艺玲, 傅懋毅. 人工林林下植被的研究现状. 林业科学研究, 2002, 15(6): 727-733. |