 2014, Vol. 34
2014, Vol. 34文章信息
- 赵明伟, 岳天祥, 孙晓芳, 赵娜
- ZHAO Mingwei, YUE Tianxiang, SUN Xiaofang, ZHAO Na
- 基于草地综合顺序分类系统(IOCSG)的中国北方草地地上生物量高精度模拟
- High accuracy simulation of aboveground biomass in Northern China based on IOCSG
- 生态学报, 2014, 34(17): 4891-4899
- Acta Ecologica Sinica, 2014, 34(17): 4891-4899
- http://dx.doi.org/10.5846/stxb201301050031
-
文章历史
- 收稿日期:2013-1-5
- 网络出版日期:2014-3-5






2. 中国科学院大学, 北京 100049;
3. 曲阜师范大学地理与旅游学院, 日照 276800
2. University of Chinese Academy of Sciences, Beijing 100049, China;
3. School of Geography and Tourism, Qufu Normal University, Rizhao 276800, China
草地生态系统是陆地植被中最重要、分布最广泛的生态系统类型之一,在全球碳循环和气候调节中起重要作用[1]。草地生物量的估算具有重要意义,一方面是因为生物量是生态系统碳库的重要组成部分[2],准确估算草地生物量对于准确评估草地生态系统的碳源汇功能具有重要意义[3, 4, 5]。另一方面,我国拥有非常丰富的草地资源,分布范围占国土面积1/3以上[6],因而准确估计我国草地生态系统的生物量及其分布,对于发展畜牧业、维持生物多样性、保持水土和维护生态系统平衡具有重要意义[7, 8, 9]。
当前估算区域尺度上的草地生物量主要是采用遥感技术,根据地面实测样点草地生物量与遥感指数(主要是各种植被指数)之间的回归关系,进而推测区域尺度上的草地生物量[10, 11, 12, 13, 14]。这种方法可以较容易估算大尺度上草地生物量,但是由于遥感自身以及植被反射等问题,基于遥感植被指数的草地生物量计算仍然存在较大的不确定性[15, 16, 17, 18]。这种不确定性产生的原因可能在于遥感植被指数只是草地生物量的一种外在反映,而不是决定草地生物量的内在影响因素,这也导致了只能估算当前或过去的草地生物量,而难以对草地生物量未来的变化趋势做出有效预测。近年来相关学者开始逐渐重视草地生物量的内在影响因素分析,例如地形因子[19, 20, 21]、年均温、相对湿度、日照时数等水热因子[22, 23, 24]。这些研究考虑了不同草地生物量的影响因素,但是一方面这些研究主要侧重于定性分析,缺乏结合影响因素的定量生物量模型构建,另一方面,这些研究没有考虑到不同区域草地生物量影响因素的差异性,而是对所研究的区域统一做相关性分析。
为了分析草地生物量的内在影响因素,并顾及这种影响因素的区域差异性,本研究首先依据草地综合顺序分类系统将所研究的区域划分为不同的潜在植被类型,将每种潜在植被类型区域内的水热条件控制在同一个水平,然后进一步分析各区域的草地生物量的内在影响因素,最后借助HASM理论模拟了研究区域的草地生物量的空间分布情况。
1 研究方法 1.1 研究区概况与数据采集根据草地生物量实测点的分布划定的研究区域如图(1)所示,包括内蒙古中部大部分区域、河北省北部以及东北三省(黑龙江,吉林和辽宁)的西部地区。经度范围为106.5°—125.5°E,纬度范围37.5°—49.5°N。该区域年平均温度变化范围为-1.8°—9.1°,年均降雨量变化范围为146—511 mm,主要植被类型为温性荒漠草原、温性草原和温性草甸草原。
本文的研究数据主要包括草地生物量实测数据和气候数据。其中气候数据为研究区域内部及周围共计104个气象站点的观测数据(1990—2000年,数据来源:http: // cdc.cma.gov.cn /home.do),主要包括月平均温度和月平均降雨量。草地生物量数据来自中国农业科学院北方样地考察数据,采样时间为1992—1994年[25]。生物量的测定方法为在草地生物量最高的8—9月,沿拟定的调查路线,采用收割法进行测定,每个样地内根据地形和土壤条件、草地组成和分布格局等因素设置5—10个代表样方,其中草本植物样方尺寸为1 m×1 m,半灌木或高大草本样方尺寸为2 m×2 m。调查数据记录了各样地的经纬度信息、植被类型、地上生物量最大时期的植被高度、植被盖度和地上生物量,收割草本植物地上部分,进行风干称重。根据本研究的需要舍弃了部分样点数据(主要是包含灌木的样点以及异常值点),最后保留了共计165个样点的数据(图 1)。

|
| 图 1 草地生物量实测样点分布 Fig. 1 Sample stations in the study area |
草地综合顺序分类系统(Integrated Orderly Classification system of Grassland,IOCSG)最初由任继周等提出[26],随后在草地类型划分、潜在草地生产力估计中得到广泛应用[27, 28]。草地综合顺序分类法根据气候特征来划分草原植被类型,这是因为地带性的生物气候条件是生物立地条件本质的体现,并且在一定的历史时期相对稳定,因此水热状况作为生物气候条件的核心,也被用来区别草原植被类型。任继周等最初将我国全年积温(>0℃)分为8个级别,将湿润度分为6个级别,因此全国共计48种类型[26]。后来又作了修正,将全年积温分为7个级别,将湿润度分为6个级别,因此全国共计42种类型[27]。本研究采用修正后的分类方案,其中本文所采用的实测数据大部分集中于最后分类结果中的4种,分别是微温干旱温带半荒漠类、微温微干温带典型草原类、微温微润草甸草原类和微温湿润森林草原类(表 1)。
根据IOCSG理论,确定潜在植被类型的依据是年积温(>0℃)和湿润度。统计每月的温度可计算年积温(>0℃)记为 ∑θ ,然后再根据年平均降水量计算湿润度:

式中, k为湿润度,R为年平均降水量,0.1为模型调整参数。
林慧龙等根据水热综合条件,建立了以年积温和湿润度为指标的草地植被生产力估算模型[29],该模型由周广胜等建立的植物净第一性生产力模型[30]推导得出,由于该模型仅仅使用相对较容易获取的年积温和湿润度作为指标,因此被很多学者采用[31, 32]。对于本研究而言,因为它综合考虑了水热因素,因此计算潜在植被NPP的目的是将其作为一个草地植被生物量的影响因子。估计NPP的基本公式为:
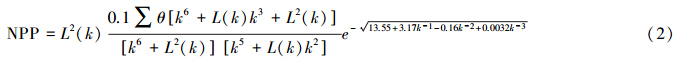
式中, L(k)=0.58802k3+0.50698k2-0.0257081k+0.0005163874 , k 为湿润度。这里需要指出的是,对于上式中 L(k) 的计算,赵军等计算公式的 k 的系数为0.257081[32].这样计算得出的NPP在 k 值较小的地区会出现负值,追溯建立该模型的文献[29],发现这其实是作者当时的个人疏忽造成的错误,作者的推导过程没有问题,但是在最后却将系一次项的系数记为0.257081,本文对此处错误作了更正。
1.3 高精度曲面模拟方法高精度曲面建模方法(HASM)是近年来针对困扰曲面建模的误差问题和多尺度问题发展起来的一种基于微分几何学曲面理论的曲面建模方法,已经成功运用于土壤插值领域、气候模拟和建立DEM等领域[33, 34, 35, 36]。
根据曲面论基本定理,一个空间曲面完全由其第一类基本量和第二类基本量决定,其中曲面第一类基本量、第二类基本量需满足如下的高斯方程:

式中, E , G 为曲面第一类基本量; L , M , N 为曲面第二类基本量, Γ111 , Γ211 , Γ122 , Γ222 , Γ112 , Γ212 称为曲面的第二类克里斯托弗尔变量。曲面第一类基本量、第二类基本量以及第二类克里斯托弗尔变量的计算公式可参见参考文献[33]。
HASM以上述高斯方程为基础,首先将空间曲面离散格网化,根据曲面驱动场建立空间曲面上的差分方程组,然后以空间实测点信息作为精度控制条件,逐步迭代求解,最后使模拟曲面最为接近真实曲面,达到最优。
首先在各潜在植被类型区建立草地生物量与其影响因素的回归模型,然后以所建模型计算得到的研究区域生物量作为驱动场,以实测点草地生物量作为精度控制条件,建立HASM的草地生物量的高精度模拟模型:
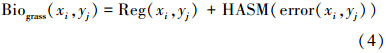
式中, Biograss(xi,yj) 表示空间位置 (xi,yj) 的草地生物量值, Reg(xi,yj) 该位置上草地生物量的回归值, HASM(error(xi,yj)) 代表由HASM模型模拟的相应空间位置上的生物量残差值。为了对比分析该方法的估算精度,本文选择两个对照模型,第一个是指数函数模型,其中自变量是NDVI最大值;第二个对照模型是HASM-GB模型,模型相关说明可参见文献[37]。
2 结果和分析 2.1 潜在植被类型划分与潜在NPP分析潜在植被类型的划分需要计算研究区域的年积温(>0℃)和湿润度,而根据式(1)湿润度的计算需要研究区域的年平均降雨量,因而需要对研究区域内气象观测站点的气候数据(月平均温度及年降雨量)进行空间插值得到气候变量在区域中的分布。根据前人的研究结果,温度的主要影响因素是高程和纬度,降水的主要影响因素则是经度和坡向[38]。首先根据研究区域内部及其周围共计104个气象站点2001—2010年的观测数据,分别建立温度和降水与各自影响因素之间的回归关系,得到研究区域的气温(每月平均)和降水(年平均)的趋势面,然后对观测值与趋势值之间的残差进行克里格插值,得到研究区域每月的温度和年平均降水量。
得到每月的温度后则可计算年积温(>0℃)记为 ∑θ ,然后再根据公式(1)计算湿润度。计算得到的研究区域的年积温和湿润度如图 2和图 3所示。计算出积温和湿润度以后,依据积温和湿润度的不同取值范围(表 1),可以确定研究区域的潜在植被类型(图 4)。可以看出,由于4种潜在植被类型区域的年积温处于同一个水平,因此湿润度指标的大小决定了潜在植被类型的差异,四种潜在植被类型的空间分布与湿润度的分布格局基本一致。具体而言,毛乌素沙地、鄂尔多斯高原及内蒙古高原北部属于微温干旱温带半荒漠类,阴山南侧、大兴安岭山脉两侧(浑善达克沙地和科尔沁沙地)及呼伦贝尔高原西部属于微温微干温带典型草原类,黄河沿岸、大兴安岭地区、呼伦贝尔高原东部及松辽分水岭北部和东部属于微温微润草甸草原类,而研究区域的东部边缘靠近西辽河及嫩江流域,属于微温湿润森林草原类。

|
| 图 2 研究区域的年积温 Fig. 2 Thermal >0℃ annual cumulative temperature |

|
| 图 3 研究区域的湿润度 Fig. 3 Humidity of the study area |

|
| 图 4 研究区域的潜在植被类型划分 Fig. 4 Distribution map for grassland classes of the study area |
基于修正后的NPP估算模型,计算得到研究样区的NPP值如图 5所示。可以看出,研究区域的潜在NPP估值从西向东NPP估值逐渐增加,并且存在变化明显的界线,变化趋势与潜在植被类型对应良好。同时,对比研究区域的年积温和湿润度(图 2,图 3)可以看出,潜在NPP值与湿润度具有更高的相似度,两者的在空间中的变化趋势基本一致,说明湿润度是影响潜在NPP值的主要影响因素。

|
| 图 5 研究样区的潜在NPP估值 Fig. 5 NPP estimation of the study area |
将年积温、湿润度和潜在NPP值作为草地生物量的影响因素,此外还考虑了地形因素,主要是坡度和坡向,土质因素主要是,表层土壤沙砾含量和表层土壤粘粒含量(数据来源:http:// globalchange.bnu.edu.cn/research/soil)。分析不同潜在植被类型区的草地生物量与上述影响因素的关系,结果如图 6所示。

|
| 图 6 不同潜在植被类型区域草地生物量影响因素分析 Fig. 6 Impact factors analysis of biomass for different grassland classes |
可以看出,对于研究样区的四种潜在植被类型,微温干旱温带半荒漠类中的草地生物量与年积温呈正相关,两者存在近似的线性关系,表明在该区域年积温的增加会增加草地生物量。微温微干典型草原类中的草地生物量与表层土壤粘粒含量呈正相关,两者的关系可以用二次多项式近似模拟,表明该区域在水热条件近似一致的情况下,土质因素是影响草地生物量的主要因素。微温微润草甸草原类和微温湿润草原类中的草地生物量则均随各自类型区域的潜在NPP值的变化呈现出先减小后增大的趋势,反映了水热因素对该两类潜在植被类型区域的草地生物量的综合影响。
对于四类潜在植被类型区域,根据各自区域的草地生物量实测值与各自影响因素的所建立的回归方程,统计了每种区域的回归误差(表 2)。为了对比分析该方法的估算精度,本文将精度分析结果与孙晓芳[37]的结果作对比,其中表 2中的对照模型1是指数函数模型,对照模型2是HASM-GB模型。可以看出,4种潜在植被类型区域分别建立的回归模型计算结果的平均误差和平均误差绝对值均小于对照模型1和对照模型2;均方根误差小于对照模型1,但大于对照模型2;4个回归关系式的相关系数均优于两个对照模型。因此,根据潜在植被类型分别建立的草地生物量的回归模型优于其它模型。
| 模拟方法 Simulation model | 平均误差 Mean error | 平均误差绝对值 Absolute mean error | 均方根误差 Root mean square error | 相关系数 Correlation coefficient |
| 潜在植被类型1 Potential vegetation type 1 | 0.1012 | 25.9316 | 31.9836 | 0.8060 |
| 潜在植被类型2 Potential vegetation type 2 | 0.0041 | 29.0267 | 33.9251 | 0.7086 |
| 潜在植被类型3 Potential vegetation type 3 | -0.0902 | 38.4635 | 44.4029 | 0.7454 |
| 潜在植被类型4 Potential vegetation type 4 | -0.3896 | 66.0154 | 87.0799 | 0.8132 |
| 对照模型1Contrast model 1 | 45.62 | 101.33 | 129.31 | 0.45 |
| 对照模型2Contrast model 2 | 3.5 | 72.81 | 28.03 | 0.62 |
对于本研究中的4种潜在植被类型,根据前面所建立的草地生物量与其影响因素之间的回归关系,可以建立相应区域的基于HASM的草地生物量的高精度模拟模型:
当潜在植被类型为微温干旱温带半荒漠类时:
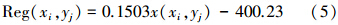
式中, x(xi,yj) 为对应空间位置的年积温。
当潜在植被类型为微温微干温带典型草原类时:

式中, x(xi,yj) 为对应空间位置的表层土壤粘粒含量百分数。
当潜在植被类型为微温微润草甸草原类时:
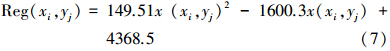
式中, x(xi,yj) 为对应空间位置的潜在NPP估值。
当潜在植被类型为微温湿润森林草原类时:
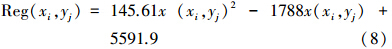
式中, x(xi,yj) 为对应空间位置的潜在NPP估值。
对于每一种潜在植被类型区域,根据回归模型计算的生物量趋势面值与运用HASM模型计算的残差值相加便得到每种潜在植被类型区域的生物量,然后将4个区域进行综合便得到本文研究区域的草地生物量高精度模拟分布(图 7)。根据计算结果,4种潜在植被类型区域所对应的草地生物量密度分别是:微温干旱温带半荒漠类为76.62g/m2,微温微干温带典型草原类为110.94 g/m2,微温微润草甸草原类为142.69 g/m2,微温湿润森林草原类为184.40 g/m2。由于四种潜在植被类型对应的湿润度是递增的,因此生物量密度同样呈现递增变化。

|
| 图 7 研究区域草地生物量高精度模拟 Fig. 7 High accuracy biomass simulation of grassland in the study area |
当前大区域尺度上草地生物量的估算集中于根据遥感植被指数进行计算,但是由于卫星遥感本身及植被反射等各种问题,基于遥感数据计算的植被指数受到很多因素的干扰而产生一定的不确定性,此外,基于植被指数的草地生物量估算方法难以实现对未来草地生物量的有效预估计。针对上述问题,本研究尝试构建不依赖于遥感指数,而是依据相对稳定的气候、土壤质地等因素构建草地生物量估算模型。水热条件是影响草地植被立地生长的核心因素,因此本研究首先采用草地植被综合顺序分类法(IOCSG)依据年积温和湿润度指标将研究区域划分为四种潜在植被类型:微温干旱温带半荒漠类、微温微干温带典型草原类、微温微润草甸草原类和微温湿润森林草原类。在此基础上对每一种潜在植被类型区域,分析草地生物量的主要影响因素,结果表明微温干旱温带半荒漠类的草地生物量与年积温存在较好的线性关系,微温微干温带典型草原类的草地生物量可以用表层土壤粘粒含量的二次多项式来模拟,后两种潜在植被类型的草地生物量则随着潜在NPP的变化呈现先减小后增大的变化趋势。然后对于每种潜在植被类型区,根据草地生物量与其影响因素的关系生成各个区域的草地生物量趋势面,最后根据HASM理论模拟生成的趋势面与地面实测生物量之间的残差分布并最终得到研究区域的草地生物量高精度模拟。最后的模拟结果显示,上述四类潜在植被类型区域的草地生物量密度值分别是:76.62、110.94 、142.69 、184.40 g/m2。
将计算结果与马文红等[39]提供的实测数据进行对比,其研究的草原类型为荒漠草原、典型草原和草甸草原,其中荒漠草原对应本研究的微温干旱温带半荒漠类,典型草原对应本研究的微温微干温带典型草原类和微温微润草甸草原类,而草甸草原对应本研究的微温湿润森林草原类。其3种草原类型的草地生物量实测平均值分别为56.6,133.4,196.7 g/m2,可以发现与本研究得出的结果趋势相同,数值上差别也不大,表明本研究得到的区域尺度上的草地地上生物量具有较高的精度。
由于实测数据的限制,本研究的研究区域主要集中在内蒙古及其与其他省相邻的边界地区。中国的草地分布范围非常广泛,从东北平原,越过大兴安岭,经辽阔的内蒙古高原,再经鄂尔多斯高原、黄土高原,一直到青藏高原,绵延约4500km,其分布范围占国土面积1/3以上,在不同区域草地生物量的影响因素可能不尽相同,因此,下一步的工作重点一方面是根据全国范围内的草地生物量实测数据,分析全国范围内各个潜在植被类型区域的草地生物量的影响因素,并在此基础上实现全国范围的草地生物量高精度模拟。另一方面,根据中国大陆气候未来情景的变化趋势分析模拟中国大陆草地生态系统生物量的未来变化趋势,是今后工作的重点。
| [1] | Hall D O, Scurlock J M O. Climate change and productivity of natural grasslands. Annals of Botany-London, 1991, 67(S1): 49-55. |
| [2] | Schlesinger W H. Carbon balance in terrestrial detritus.Annual Review of Ecology and Systematics, 1977, 8(1): 51-81. |
| [3] | Sala O E, Lauenroth W K, Burke I. Carbon budgets of temperate grasslands and the effects of global change//Breymeyer A I, Hall D O, Melillo J M, eds. Global Change: Effects on Coniferous Forests and Grasslands. London: John Wiley & Sons Ltd, 1996. |
| [4] | Scurlock J M O, Johnson K, Olson R J. Estimating net primary productivity from grassland biomass dynamics measurements. Global Change Biology, 2002, 8(8): 736-753. |
| [5] | Parton W J, Scurlock J M O, Ojima D S, Schimel D S, Hall D O. Impact of climate change on grassland production and soil carbon worldwide. Global Change Biology, 1995, 1(1): 13-22. |
| [6] | The Department of Animal Husbandry and Veterinary and the General Station of Animal Husbandry and Veterinary of the Ministry of Agriculture, P. R. China. Rangeland Resources of China. Beijing: Chinese Science and Technology Press, 1996. |
| [7] | Xie G D, Zhang Y L, Lu C X, Zheng D, Cheng S K. Study on valuation of rangeland ecosystem services of China. Journal of Natural Resources, 2001, 16(1): 47-53. |
| [8] | Hudak A T, Brockett B H. Mapping fire scars in a southern African savannah using Landsat imagery. International Journal of Remote Sensing, 2004, 25(16): 3231-3243. |
| [9] | Mutanga O, Skidmore A K. Integrating imaging spectroscopy and neural networks to map grass quality in the Kruger National Park, South Africa. Remote Sensing of Environment, 2004, 90(1): 104-115. |
| [10] | Piao S L, Fang J Y, Zhou L M, Tan K, Tao S. Changes in biomass carbon stocks in China's grasslands between 1982 and 1999. Global Biogeochemical Cycles, 2007, 21(2): GB2002. |
| [11] | Fang J Y, Guo Z D, Piao S L, Chen A P. Terrestrial vegetation carbon sinks in China, 1981-2000. Science in China Series D: Earth Sciences, 2007, 50(9): 1341-1350. |
| [12] | Xie Y C, Sha Z Y, Yu M, Bai Y F, Zhang L. A comparison of two models with Landsat data for estimating above ground grassland biomass in Inner Mongolia, China. Ecological Modelling, 2009, 220(15): 1810-1818. |
| [13] | Xin X P, Zhnag B H, Li G, Zhang H B, Chen B R, Yang G X. Variation in spatial pattern of grassland biomass in China from 1982 to 2003. Journal of Natural Resources, 2009, 24(9): 1583-1592. |
| [14] | Jin Y X, Xu B, Yang X C, Li J Y, Wang D L, Ma H L. Remote sensing dynamic estimation of grass production in Xilinguole, Inner Mongolia. Scientia Sinica(Vitae), 2011, 45(12): 1185-1195. |
| [15] | Wen J, Wang J M. A modified soil-adjusted vegetation index obtained from satellite remote sensing data. Climatic and Environmental Research, 1997, 2(3): 302-309. |
| [16] | Kogan F N, Zhu X. Evolution of long-term errors in NDVI time series: 1985-1999. Advances in Space Research, 2001, 28(1): 149-153. |
| [17] | Trishchenko A P, Cihlar J, Li Z Q. Effects of spectral response function on surface reflectance and NDVI measured with moderate resolution satellite sensors. Remote Sensing of Environment, 2002, 81(1): 1-18. |
| [18] | Zhang Y N, Niu J M, Zhang Q, Yang Y, Dong J J. A discussion on applications of vegetation index for estimating aboveground biomass of typical steppe. Acta Prataculturae Sinica, 2012, 21(1): 229-238. |
| [19] | Perelman S B, Leon R J C, Oesterheld M. Cross-scale vegetation patterns of Flooding Pampa grasslands. Journal of Ecology, 2001, 89(4): 562-577. |
| [20] | López I F, Lambert M G, Mackay A D, Valentine I. The influence of topography and pasture management on soil characteristics and herbage accumulation in hill pasture in the North Island of New Zealand. Plant and Soil, 2003, 255(2): 421-434. |
| [21] | Bennie J, Hill M O, Baxter R, Huntley B. Influence of slope and aspect on long-term vegetation change in British chalk grasslands. Journal of Ecology, 2006, 94(2): 355-368. |
| [22] | Sitch S, Smith B, Prentice I C, Arneth A, Bondeau A, Cramer W, Kaplan J O, Levis S, Lucht W, Sykes M T, Thonicke K, Venevsky S. Evaluation of ecosystem dynamics, plant geography and terrestrial carbon cycling in the LPJ dynamic global vegetation model. Global Change Biology, 2003, 9(2): 161-185. |
| [23] | Ni J. Estimating net primary productivity of grasslands from field biomass measurements in temperate northern China. Plant Ecology, 2004, 174(2): 217-234. |
| [24] | Hu Z M, Fan J W, Zhong H P, Yu G R. Spatiotemporal dynamics of aboveground primary productivity along a precipitation gradient in Chinese temperate grassland. Science in China (Series D), 2006, 50(5): 754-764. |
| [25] | Institute of Grassland Research of CAS. Reserch on Dynamics Monitoring of Grazing Ecosystem in the North of China: The Dataset of Dynamics Monitoring of Grazing Ecosystem in the North China. Hohhot: Inner Mongolia Press, 1996. |
| [26] | Ren J Z, Hu Z Z, Mou X D, Zhang P J. Comprehensive and sequential classification system of grasslands and its significance analysis. Chinese Journal of Grassland, 1980, 1(1): 12-24. |
| [27] | Ren J Z, H u Z Z, Zhao J, Zhang D G, Hou F J, Lin H L, Mu X D. A grassland classification system and its application in China. The Rangeland Journal, 2008, 30(2): 199-2091. |
| [28] | Zhang M L, Chen Q G, Jiang W L, Liu X N. An improved CASA model based on comprehensive and sequential classification system of grasslands. Chinese Journal of Grassland, 2011, 33(4): 5-11. |
| [29] | Lin H L, Wang J, Xu Z. Study on grassland net primary productivity (NPP) model using ≥0 ℃ annual average cumulative temperature(∑θ)and moisture index (K). Pratacultural Science, 2005, 22(6): 8-10. |
| [30] | Zhou G S, Zhang X S. A natural vegetation NPP model. Acta Phytoecologica Sinica, 1995, 19(3): 193-200. |
| [31] | Gong Y M, Hu Y K, Adeli M D, Li K H, Yi W, Zhang W, Wang J Y. Analysis of adaptation of a climate productivity model on alpine grassland. Acta Prataculturae Sinica, 2010, 19(2): 7-13. |
| [32] | Zhao J, Shi Y F, Wang D W. Analysis of spatial distribution features of potential vegetation NPP in Inner Mongolia based on the IOCS. Journal of Natural Resources, 2012, 27(11): 1870-1880. |
| [33] | Yue T X. Surface Modeling: High Accuracy and High Speed Methods. Beijing: CRC Press, 2011. |
| [34] | Yue T X, Wang S H. Adjustment computation of HASM: a high-accuracy and high-speed method. International Journal of Geographical Information Science, 2010, 24(11): 1725-1743. |
| [35] | Yue T X, Du Z P, Song D J, Gong Y. A new method of surface modeling and its application to DEM construction. Geomorphology, 2007, 91(1/2): 161-172. |
| [36] | Shi W J, Liu J Y, Song Y J, Du Z P, Chen G F, Yue T X. Surface modelling of soil pH. Geoderma, 2009, 150(1/2): 113-119. |
| [37] | Sun X F. High Accuracy Surface Modeling of Vegetation Aboveground Biomass in the Major Terrestrial Ecosystems . Beijing: Institute of Geographical Sciences and Natural Resources Research, Chinese Academy of Sciences. |
| [38] | Wang C L, Yue T X, Fan Z M, Zhao N, Sun X F. HASM-based climatic downscaling model over China. Journal of Geo-Information Science, 2012, 14(5): 599-610. |
| [39] | Ma W H, Fang J Y, Yang Y H, Mohammat A. Biomass carbon stocks and their changes in northern China's grasslands during 1982-2006. Science China Life Sciences, 2010, 53(7): 841-850. |
| [6] | 中华人民共和国农业部畜牧医司, 全国畜牧兽医总站. 中国草地资源. 北京: 科学出版社, 1996. |
| [13] | 辛晓平, 张保辉, 李刚, 张宏斌, 陈宝瑞, 杨桂霞. 1982-2003 年中国草地生物量时空格局变化研究. 自然资源学报, 2009, 24(09): 1583-1592. |
| [14] | 金云翔, 徐斌, 杨秀春, 李金亚, 王道龙, 马海龙. 内蒙古锡林郭勒盟草原产草量动态遥感估算. 中国科学: 生命科学, 2011, 45(12): 1185-1195. |
| [15] | 文军, 王介民. 一种由卫星遥感资料获得的修正的土壤调整植被指数. 气候与环境研究, 1997, 2(3): 302-309. |
| [18] | 张艳楠, 牛建明, 张庆, 杨艳, 董建军. 植被指数在典型草原生物量遥感估测: 应用中的问题探讨. 草业学报, 2012, 21(1): 229-238. |
| [25] | 中国农业科学院草原研究所. 中国北方草地畜牧业动态监测数据集: 中国北方草地畜牧业动态监测研究(二). 呼和浩特: 内蒙古大学出版社, 1996. |
| [26] | 任继周, 胡自治, 牟新待, 张普金. 草原的综合顺序分类法及其草原发生学意义. 中国草地学报, 1980, 1(1): 12-24. |
| [28] | 张美玲, 陈全功, 蒋文兰, 柳小妮. 基于草地综合顺序分类法的CASA模型改进. 中国草地学报, 2011, 33(4): 5-11. |
| [29] | 林慧龙, 王军, 徐震. 草地净第一性生产力与≥0℃年积温、湿润度指标间的关系. 草业科学, 2005, 22(6): 8-10. |
| [30] | 周广胜, 张新时. 自然植被净第一性生产力模型初探. 植物生态学报, 1995, 19(3): 193-200. |
| [31] | 公延明, 胡玉昆, 阿德力·麦地, 李凯辉, 尹伟, 张伟, 王吉云. 高寒草原对气候生产力模型的适用性分析. 草业学报, 2010, 19(2): 7-13. |
| [32] | 赵军, 师银芳, 王大为. 基于IOCS的内蒙古潜在植被NPP空间分布特征研究. 自然资源学报, 2012, 27(11): 1870-1880. |
| [37] | 孙晓芳. 主要陆地生态系统植被地上生物量高精度曲面建模 [D]. 北京: 中国科学院研究生院, 2012. |
| [38] | 王晨亮, 岳天祥, 范泽孟, 赵娜, 孙晓芳. 高精度曲面建模的中国气候降尺度模型. 地球信息科学学报, 2012, 14(5): 599-610. |