 2014, Vol. 34
2014, Vol. 34文章信息
- 刘玉莉, 江洪, 周国模, 陈云飞, 孙成, 杨爽
- LIU Yuli, JIANG Hong, ZHOU Guomo, CHEN Yunfei, SUN Cheng, YANG Shuang
- 安吉毛竹林水汽通量变化特征及其与环境因子的关系
- Water vapor flux variation characteristic and the relationship with its environment factors in phyllostachys edulis forest in Anji
- 生态学报, 2014, 34(17): 4900-4909
- Acta Ecologica Sinica, 2014, 34(17): 4900-4909
- http://dx.doi.org/10.5846/stxb201301050025
-
文章历史
- 收稿日期:2013-1-5
- 网络出版日期:2014-3-5






2. 南京大学国际地球系统科学研究所, 南京 210093
2. International Institute for Earth System Science, Nanjing University, Nanjing 210093, China
毛竹phyllostachys edulis是禾本科Poaceae竹亚科刚竹属竹种,单轴散生型,多年生常绿植物,根系稠密集中,生长周期短,产量高。毛竹林是我国重要的森林资源之一,长江以南,分布着世界上85%的毛竹,且大多生长在400—800 m的丘陵、低山山麓地带。研究区安吉县更素有“毛竹之乡”的美誉。近年来,对毛竹的光合生理[1, 2, 3]、土壤呼吸特征[4]、以及固碳能力[5, 6]研究的相对较多,但是,迄今为止仍未见与毛竹林水汽通量相关的研究报道。水汽通量是生态系统水循环过程的一个重要特征参数,又称水汽输送量,是单位时间内通过单位面积的水汽量。陆地-大气系统的水蒸气输送是水循环的一个环节,潜热输送的载体,能量平衡的重要影响因子。森林水汽通量主要指地面或水面的蒸发通量、植被冠层截留降水的蒸发通量和植物的蒸腾通量三者之和,是森林植被水分状况的重要指标,生态系统能量闭合的重要影响因素,影响区域和全球气候的重要因子。尤其是地表植被的蒸散通量一直被作为影响全球气候的重要因素之一[7, 8],发展至今,随着通量观测系统及其技术的出现,利用涡度相关技术研究蒸散[9, 10]日益受到学者们的广泛关注。
目前,在全球范围内,微气象学的涡度相关法已应用在陆地生态系统物质和能量交换观测,并且取得良好效果。这种方法也成为通量观测网络FLUXNET(Ameri-Flux、Euro-Flux、Asia-Flux、China-Flux) 的标准观测方法[11, 12, 13]。本研究以浙江省安吉县山川乡毛竹林生态系统为研究对象,利用涡度相关观测技术研究了毛竹林2011年全年水汽通量的动态变化特征,分析了其与净辐射、饱和水汽压等环境因子之间的关系,为进一步研究毛竹林生态系统的水汽收支状况提供一定的科学依据,也为当地毛竹经营管理的可持续发展提供一定的理论参考。
1 材料与方法 1.1 试验地概况试验区设在浙江省湖州市安吉县山川乡,地理位置30°28′34.5″N,119°40′25.7″E,属于亚热带季风气候类型。气候特点:季风显著,四季分明,雨热同期,有梅雨季节,空气湿润。年平均气温16.6 ℃,年日照时数1613—2430 h,1月份温度最低,平均气温-0.4—5.5 ℃,7月份温度最高,平均气温24.4—30.8 ℃,年降水量761—1780 mm,月平均相对湿度在70%以上。安吉县山川乡森林面积为4251 hm2,其中竹林面积为2155 hm2,占森林总面积的50.7%,毛竹林面积1693 hm2,占竹林总面积的78.6%。毛竹林通量观测塔位于海拔380 m处,(土壤类型为黄壤、黄红壤,下垫面坡度2.5—14°,坡向为北偏东8°),周围1000 m范围之内的植物类型以毛竹为主。试验区内的毛竹林为人工纯林,林分密度为4500株 /hm2,毛竹胸径12—18 cm,高度13—20 m,枝下高10—17 m,冠幅3.0 m×2.5 m,盖度90%,郁闭度0.9,毛竹节间短,壁厚,出笋期为3—5月,且连年出笋能力较强,平均每年产量约为1000—1500 kg/hm2,林下有极少灌木和草本。
1.2 观测仪器竹林中建有高40 m的微气象观测塔,开路涡度相关系统的探头安装在距地面38 m的高度,主要由三维超声风速仪(CAST3,Campbell Inc.,USA) 和开路CO2/H2O分析仪(Li-7500,LiCor Inc.,USA) 组成,其原始采样频率为10 Hz,数据采集器(CR1000,Campbell Inc.,USA) 存储所有数据,可在线计算并存储30 min的CO2通量(carbon flux,FC)、摩擦风速(Ustar)、潜热通量(latent heat flux,LE)和显热通量(sensible heat flux,HS)等结果。
常规气象观测系统主要包括3层风速(010C,metone,USA)、3层大气温度和湿度(HMP45C,Vaisala,Helsinki,Finland),安装高度分别为1 、7 m 和38 m。2个SI-111红外温度分别置于2、23 m,采集地表和冠层的温度;安装在38 m的高度的净辐射仪(CNR4,Kipp&Zonen)传感器,分别用来采集上行和下行的长波/短波辐射以及净辐射的数据。常规气象观测系统的数据采样频率为0.5 Hz,该系统通过数据采集器(CR1000,Campbell Inc,USA) 每30 min自动记录平均风速、气压、温度和净辐射等常规气象信息。试验样地内设有雨量筒,用于记录降水量信息。
1.3 研究方法 1.3.1 计算公式水汽通量(E) 通过实时测定的垂直风速与其浓度的协方差来求得。采用的公式为:
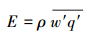
式中,ρ代表干空气密度,q代表比湿脉动,w代表垂直风速;横线表示一段时间内的平均值;撇号表示脉动。并规定若气体由大气圈进入生态系统,通量符号为负,若气体由生态系统进入大气圈,则通量符号为正[14]。
1.3.2 数据处理该试验采用的数据为通量观测的30 min平均值。数据处理采用目前普遍采用的比较成熟的方法,主要包括2次坐标旋转来矫正地形以及观测仪器的不水平,并使垂直方向的风速平均值为0,水平方向的风速和主导风向一致,且剔除由于恶劣天气(有降水)、湍流不充分等导致的不合理数据,对于打雷、仪器故障等原因导致的缺失数据采取如下方法插补:其中≤2 h的用平均值来插补,即用平均日变化法(MDV)插补缺失的数据[15],对于缺失的数据采用相邻几天相同时刻的平均值来进行插补,此方法首先要确定平均时段的长度,另有研究表明白天取14 d、夜间取7 d的平均时间长度所得结果的偏差是最小的[16];>2 h的用其与净辐射的方程插补。
2 结果与分析 2.1 全年水汽通量各月平均日变化特征对安吉毛竹林2011年全年水汽通量数据进行统计,得到逐日逐半小时的水汽通量数据,按月将每天同时刻的水汽通量求平均值,计算当月平均日变化,结果如图 1所示。

|
| 图 1 2011年安吉毛竹林水汽通量各月平均日变化分布 Fig. 1 Mean diurnal change distribution of monthly water vapor flux in phyllostachys edulis forest of Anji country(2011) |
图 1显示,毛竹林全年水汽通量基本为正值,均呈单峰型变化趋势,表明水汽输送方向是由毛竹林生态系统向大气输送,毛竹林是水汽源。各月最大值均在12:00—14:00出现,呈现一定规律性,是由于该段时间太阳辐射较强,温度较高,湍流通量较大,植物蒸腾和地表蒸发比较旺盛;而夜间水汽通量几乎为0,原因是夜间温度较低,光线较弱,土壤蒸发和叶片蒸腾极其微弱。同时净辐射为负值,湍流通量很小。6:00—8:00逐渐升高,至最高点后逐渐降低,17:00—19:00趋近于0,波动较为平缓,且基本保持稳定。一昼夜内,生态系统的温度、热量、水分等气象因子都发生明显变化,同时植物生理活动也受昼夜生物节律的调节,因此植被蒸散有明显的日变化特征。分析各月水汽通量的变化时刻与当时日出、日落时间,凌晨到日出前植物生理活动很弱,稳定地处于一天最低水平;日出后植物开始进行光合作用,随着光照的增强,植物的生理活动越来越活跃,叶片蒸腾作用旺盛,温度升高促进土壤蒸发,蒸散迅速增加;午后1—2 h蒸散达到一天中最大值;此后开始逐步回落,到夜间蒸散处于较为稳定的低水平。
3—5月曲线较其他月份的波动大,日变化幅度较大,原因是此时气候复杂多变,温度变化幅度大,进一步影响毛竹生态系统内的地表蒸发和植被蒸腾,从而导致生态系统的水汽通量日变化幅度较大。
2011年各月水汽通量最大值在0.02—0.12 g m-2 s-1之间,不同季节差异很明显。峰值基本在12:00—14:00出现。7月水汽通量值(0.1116 g m-2 s-1)明显大于其余月份,原因是该月的光照较强,温度较高,而降水量仅为192.7 mm,植物生长受抑制,蒸腾旺盛,地表植被及土壤蒸发较大。 12月最低(0.0209 g m-2 s-1),这也与当月的气候条件以及下垫面性质相吻合。
毛竹林全年水汽通量最小值基本为-0.0024—0.0166 g m-2 s-1,且均集中在夜间或凌晨,只是各月出现的时间点不同。6、8月的夜间水汽通量明显大于其它月份,其中8月(0.0166 g m-2 s-1)最大,是由于这两个月处于夏季,降水较多,空气湿度大,气温高,白天云层较厚,吸收光照热量,致使毛竹林温度变化幅度相对较小,水汽通量无明显变化;夜间云层释放热量,致使毛竹林内部温度较其余月份稍高,夜间水汽通量较之稍高,18:00—20:00气温下降,水汽通量下降,趋近于0。
2.2 全年水汽通量季节变化特征通过对全年数据的统计,得到安吉毛竹林生态系统的全年逐日逐半小时的水汽通量数据,按季节计算平均日变化,结果如图 2所示。

|
| 图 2 2011年安吉毛竹林水汽通量各季节平均日变化特征 Fig. 2 Mean diurnal change distribution of quarterly water vapor flux in phyllostachys edulis forest of Anji(2011) |
图 2显示,毛竹林夏季水汽通量日变化规律性较强,变化曲线为单峰,曲线平滑,18:00—次日06:00水汽通量维持在0.0075—0.0251 g m-2 s-1之间,06:00—12:00逐渐升高,在12:00左右达到最大值后逐渐降低,在18:00左右趋于稳定。秋季的变化规律性次之,变化曲线为单峰,曲线相对平缓,变化规律与之相似。而冬春两季的日变化与夏秋季相比,变化规律性较差,日变化幅度较大,表现为曲线波动较大,不够平滑,曲线除一个顶峰以外,还有若干小峰存在。分析夏季水汽通量日变化规律较强的原因是夏季为雨季,有持续阴天降水的气候现象,空气湿度大,气温较高,较其他3个季节加强了地表蒸发和植被蒸腾作用,进而导致水汽通量的增大。
由于受当地气候影响,试验区2011年水汽通量季节变化特征为:夏季特征最明显,且高于其他各季节,春秋季变化特征相似,峰值相同,冬季变化相对较复杂,有若干个峰值,曲线波动较多。
2.3 降水量与蒸散量蒸散作用是指地表蒸发作用与植物蒸腾作用的总和,为地表到大气的能量转移贡献了75%,大气中 的垂直水交换主要由太阳能通过蒸散作用来驱动,换言之,蒸散作用是水循环的一个重要因子,地球能量收支中的一个非常重要过程[17]。蒸散量是生态系统内土壤蒸发和植被蒸腾的总耗水量,是全年水汽通量的总和。主要受蒸发势、土壤供水状况、植被状况等因素影响。
通过雨量筒测得2011年各月降水量,计算各月水汽通量的总和(图 3)。

|
| 图 3 2011年安吉毛竹林蒸散量与降水量对比 Fig. 3 The contrast between evapotranspiration and Precipitation in phyllostachys edulis forest of AnJi(2011) |
图 3显示,毛竹林2、4、5月和11月的蒸散量稍大于降水量,12月二者相差不大,其余各月蒸散量均小于降水量,6月降水量远远大于蒸散量,是由于在蒸散量相对稳定的情况下,进入夏季,6月份已是梅雨期,降水大幅度增多。各季节降水量、蒸散量及其占全年的比例如表 1所示。
| 季节 Season | 月份 Month | 降水Precipitation | 蒸散Evapotranspiration | ||
| 降水量/mm Precipitation | 比例/% Proportion | 蒸散量/mm Evapotranspiration | 比例/% Proportion | ||
| 冬季Winter | 12,1,2 | 93.00 | 6.04 | 81.20 | 10.91 |
| 春季Spring | 3,4,5 | 174.70 | 11.32 | 185.67 | 24.93 |
| 夏季Summer | 6,7,8 | 1062.20 | 68.84 | 315.55 | 42.37 |
| 秋季Autumn | 9,10,11 | 213.20 | 13.80 | 162.31 | 21.79 |
| 全年All the year | 1543.10 | 744.73 | |||
毛竹林夏季的降水量为1062.20 mm,占全年总降水量(1543.10 mm)的68.84%。春秋季降水量相差不大,春季占全年的11.32%,秋季占13.80%,冬季最少,仅占6.04%。蒸散量方面,夏季最大315.55 mm,占全年蒸散量(744.73 mm)的42.37%,春季占24.93%,秋季占21.79%,冬季最小,占10.91%。季节尺度上,二者存在较强响应关系。毛竹林全年蒸散量(744.73 mm)占全年降水量(1543.10 mm)的48.26%,略低于实际情况,原因可能是夜间降水或露水对水汽通量的观测有较大影响,易导致低估通量值[18, 19],另外处理数据时采用的方法也可能是导致结果低于实际观测值的原因之一[14]。
表 2为目前中国通量观测系统(China-Flux)的部分站点,亚热带同气候类型,降水量大体相同,不同林型的生态系统进行比较,千烟洲人工针叶林 (Coniferous forest),主要树种有马尾松(P.massoniana) 、 湿地松( Pinusell iottii ) 、杉木(Cunninghamia lanceolata) 2004年全年蒸散量占全年降水量55.60%;广东鼎湖山针阔混交林优势树种为荷木(Schima superba)、锥栗(Castanopsis chinensis)和马尾松,年均蒸散量占同期降水量的58.68%;湖南会同杉木林多年年均蒸散量占同期降水量的70.50%;长江滩地抑螺防病林2006年蒸散量占同期降水量的48.45%;云南哀牢山亚热带季雨林年均蒸散量为1369.4 kg/m2;江苏常熟农田年均蒸散量为705.00 kg/m2;各生态系统全年蒸散量关系为:云南哀牢山季雨林最大,鼎湖山针阔混交林其次,湖南会同杉木林较之略小,都大于毛竹林,其余都小于毛竹林。表明不同植被类型之间蒸散量差异很大。一般而言,不同生态系统年蒸散量关系为森林>农田>草地。而森林中,高大的乔木林>亚乔木林>灌木林[23]。
| 站点 Site | 生态系统 Ecosystem | 气候类型 Climate type | 观测时间 Observation time | 年降水量/mm Annual precipitation | 年蒸散量/mm Annual evapotran Xspiration | 数据来源 Data sources |
| 江西千烟洲 | 人工针叶林 | 亚热带季风气候 | 2004 | 1323.60 | 736.10 | [14] |
| 广东鼎湖山 | 针阔混交林 | 南亚热带湿润季风气候 | — | 1900(年均) | 1115 (年均) | [20] |
| 湖南会同 | 杉木林 | 中亚热带湿润季风气候 | 1990—2005 | 1488(年均) | 1049(年均) | [21] |
| 长江滩地 | 抑螺防病林 | 亚热带湿润季风气候 | 2006 | 1500.63 | 727.06 | [22] |
| 云南哀牢山 | 季雨林 | 亚热带季风气候 | — | — | 1369.4(年均) | [23] |
| 浙江 | 毛竹林 | 亚热带季风气候 | 2011 | 1543.10 | 744.73 | 本文 |
| 江苏常熟 | 农田 | 亚热带季风气候 | — | — | 705.00(年均) | [24] |
通过对毛竹林2011年全年净辐射的数据分析,选取各季节典型月份(1、4、7月和10月)的逐日逐半小时净辐射的数据,按月将每天同时刻的净辐射求平均值来计算当月平均日变化,结果如图 4所示。

|
| 图 4 净辐射的日均变化 Fig. 4 Diurnal variation of mean value of net radiation |
毛竹林1、4、7月和10月的净辐射日变化表现为单峰型,日出后,净辐射向正值转变,在转折瞬间净辐射为0,之后不断增大,12:00左右达到一天的最大值后逐渐下降,日落净辐射由正值转为负值,转折瞬间为0,夜间太阳辐射为0,而地表温度较高,致使向外的辐射输出远大于收入,夜间净辐射就表现为负值。对比可知,7月份的峰型与其余3个月份不同,表现为12:00左右出现下降趋势,这与7月份的特殊天气有关,7月份为雨季,白天太阳辐射比较强,大气对流强烈,易在午后形成积雨云,有雷阵雨,从而引起净辐射的变化。这也解释了图 1中7月份水汽通量日变化曲线中在12:00左右出现下降趋势的现象。
2.5 净辐射与水汽通量在季节尺度上的相关性毛竹林全年净辐射为2604.821 MJ m-2 a-1,季节尺度上水汽通量对净辐射的响应见图 5。

|
| 图 5 各季度水汽通量对净辐射的响应拟合曲线 Fig. 5 The quarterly response of water vapor flux to net radiation(Rn) |
生态系统的净辐射是驱动植被下垫面温度变化、显热和潜热交换的能量来源,从根本上说,这些变化所需的能量皆由辐射平衡的能量转化而来[25],可见净辐射也是蒸散的驱动力。一般情况下,净辐射是白天水分蒸散的主要能量来源,因此,白天的蒸散量与净辐射是密切相关的[26]。净辐射Rn在不同季节变化不同,但水汽通量都对其有良好的响应。图 4显示(通过F检验),夏季相关系数最大,R2为0.6111,相关度最高,秋季R2为0.5295,春季R2为0.2605,冬季R2为0.0455。但相关系数最大时,R2为0.6111,在拟合方程均为一元二次多项式时,略高于千烟洲人工林晴天的R2(0.55)[14],小于长江滩地抑螺防病林晴天时拟合方程(y=0.0002x+0.026) 的 R2(0.82)[22],小于同时期植物生长季内三江平原典型沼泽湿地蒸散量拟合方程(y=0.379x+11.679)的R2(0.841)[27],表明毛竹林夏、秋季净辐射Rn和水汽通量线性关系较好;在植物生长期内,净辐射增大致使气温升高,水分蒸发速率增大。在强烈的太阳辐射作用下(太阳辐射与净辐射高度正相关),叶温升高,增大叶片水汽扩散梯度,有利于植物水分蒸腾;太阳辐射能够促使气孔导度增加,气孔开放,蒸腾作用加强,这是夏季净辐射对毛竹林水汽通量影响明显的根本原因。
R2大小说明影响水汽通量的因素是多方面的。通过F检验(P<0.01),水汽通量和净辐射的相关性极显著,水汽通量对净辐射有良好的响应性。与千烟洲人工针叶林、长江滩地抑螺防病林和三江平原的沼泽湿地的研究结果一致。而各季节相关系数的大小关系说明了各季节气候条件对其影响。春季气候相对复杂,天气变化多样,温度变幅较大,对水汽通量有一定的影响,图上各点比较分散、不集中,与吉喜斌等[28]的研究相符。冬季水汽通量普遍较低,且毛竹林冬季降水量较少,几乎与蒸散量持平,温度较低,有人为钩梢、冬笋出土,也一定程度影响植被蒸腾和地表蒸发量,故冬季水汽通量与净辐射相关性较小。
2.6 饱和水汽压差对水汽通量的影响蒸散发生在土壤、植被与大气间的水汽交换过程,土壤-大气之间和植被-大气之间的水汽浓度都为植被蒸散提供驱动力,因此水分状况直接影响蒸散过程。而表征大气水分状况的指标主要有水汽浓度、湿度、水汽压、相对湿度和饱和水汽压差,其中最能体现水汽密度差异指标的是饱和水汽压差(VPD),可代表性的反映大气对水分的“需求”,代表植被蒸散的驱动力[23]。
根据全年蒸散量情况,选取6—9月毛竹主要生长期的逐日逐半小时的、高度为林冠距地面1 m的饱和水汽压差的数据,图 6为对应时刻水汽通量对饱和水汽压差的响应。

|
| 图 6 2011年6月—9月安吉毛竹林饱和水汽压(VPD)对水汽通量的影响 Fig. 6 The influence of vapor pressure difference on water vapor flux in Phyllostachys edulis forest of AnJi(2011-06—09) |
图 6表明6—9月毛竹林水汽通量与VPD正相关性明显。主要原因:一毛竹叶片蒸腾作用与气孔开度和外界的空气水汽压差成正比关系,进一步与空气饱和水汽压差成正比关系[29];二水分蒸发也受饱和水汽压差影响,空气饱和水汽压差越大,蒸发作用越强。
图 6显示出水汽通量在不同月份对VPD的响应程度不相同。线性拟合程度上,一元二次方程拟合比指数拟合相关系数要高,通过F检验(P<0.01),水汽通量与VPD有极显著的相关性。6、7月份毛竹生长旺盛,需水量大,蒸腾作用强烈,对环境因子的耐受范围较大,水汽通量随着VPD的增大而增大,与张新建等[9]研究结论一致;8月毛竹发育进入成熟期,各种生理活动主要用来维持自身新陈代谢,当VPD增大到一定程度后,明显开始受到水分胁迫,叶片气孔更多的开始表现为闭合,通过降低蒸腾作用来保持自身水分平衡,水汽通量开始随VPD的增大而减小,结果与Takagi等[30]的一致。9月毛竹叶片开始枯萎,各种生理活动开始减弱,气孔闭合对水分蒸腾的限制作用增强,在一定范围内,水汽通量随VPD的增大而减小,表现为负相关关系。
3 结论安吉毛竹林2011年水汽通量特征的观测结果表明,全年各月各时刻的水汽通量基本均为正值,说明毛竹林是水汽源,月尺度上,各月日均变化呈单峰型,6:00—8:00开始逐渐升高,在12:00—14:00左右至最高点后逐渐降低,17:00—19:00趋近于0,波动较为平缓,基本保持稳定,呈现一定的规律性。7月全年最高,12月最低。
季节尺度上,不同季节水汽通量日变化特征不同。夏季日变化特征为典型单峰型,规律性较强,春秋季与之相似,冬季虽为单峰,但有若干小峰存在,规律性稍差,曲线波动大。
毛竹林全年蒸散量占全年降水量的48.26%。2、4、5、11月的蒸散量稍大于降水量,12月二者相差不大,其余各月蒸散量均小于降水量,6月降水量远大于蒸散量。
毛竹林全年净辐射为2604.821 MJ m-2 a-1,净辐射作为影响水汽通量的最主要因子,其大小直接影响当地毛竹林生态系统的水汽通量收支情况,毛竹林水汽通量状况与当地净辐射表现出较好的正相关性。
6—9月毛竹主要生长期内探讨水汽通量对饱和水汽压差的响应情况。在毛竹生长阶段,水汽通量随饱和水汽压差的增大而增大;发育成熟后,一定范围内,水汽通量随饱和水汽压的增大而增大,超过一定范围,饱和水汽压差的增大反而抑制了水分的蒸散。
水汽通量是生态系统水循环过程的重要特征参数,水循环的环节,潜热输送的载体,能量平衡的重要影响因子,因而一直是气象学、水文学以及生态学等重点学科的研究热点。由于涡度相关法的出现,极大促进了该项研究的进展,但其也有局限性,如数据的检验和修正等,本文只是对安吉毛竹林2011年全年的水汽通量以及净辐射数据进行的分析,数据只有1a,由于夜间水汽通量的校正方法以及适合全天数据缺失的插补方法等因素,对于研究区域水汽通量还存在一定的偶然性、局限性,因此需要进一步的研究来分析和验证。
| [1] | Wen G S, Zhang L Y, Zhang R M, Wang D J, Zhang J. Climate change response using a simulation study of photosynthetic physiology on Phyllostachys pubescens. Journal of Zhejiang A&F University, 2011, 28(4): 555-561. |
| [2] | Zhang L Y, Wen G S, Wang S J, Liu Z L. Four light-response models to estimate photosynthesis of Phyllostachys pubescens. Journal of Zhejiang A&F University, 2011, 28(2):188-193. |
| [3] | Shi J M, Guo Q R, Yang G Y. Study on the photosynthetic dynamic variation of Phyllostachys edulis. Forest Research, 2005, 18(5): 551-555. |
| [4] | Yuan H Y, Liu Y Y, Zhou G M, Yu S Q. Soil respiration and its regulating factors in the Phyllostachys edulis forest of west Tianmu Mountain. China. Acta Ecologica Sinica, 2010, 30 (17): 4590-4597. |
| [5] | Xiao F M, Fan S H, Wang S L, Guan F Y, Yu X J, Shen Z Q. Estimation of the carbon balance in Moso Bamboo and Chinese Fir plantation ecosystem. Scientla Silvae Sinicae, 2010,46 (11):59-64. |
| [6] | Zhou G M. Carbon storage, fixation and distribution in mao bamboo (Phyllostachys Pubescens) stands ecosystem [D]. Zhejiang: Zhejiang University, 2006. |
| [7] | Shukla J, Mintz Y. Influence of land-surface evapotranspiration on the Earth's Climate. Science, 1982, 215 (4539):1498-1501. |
| [8] | Irmak S, Asce M. Dynamics of nocturnal, daytime, and sum-of-hourly evapotranspiration and other surface energy fluxes over Nonstressed maize canopy. Journal of Irrigation and Drainage Engineering, 2011, 137(8):475-490. |
| [9] | Zhang X J, Jin C J, Guan D X, Wang A Z, Wu J B, Yuan F H. Long-Term eddy covariance monitoring of evapotranspiration and its environmental factors in a temperate mixed forest in Northeast China. Journal of Hydrologic Engineering, 2012, 17(9):965-974. |
| [10] | Liu D, Li J, Yu Q, Tong X J, Ouyang Z. Energy balance closure and its effects on evapotranspiration measurements with the eddy covariance technique in a cropland. Acta Ecologica Sinica, 2012, 32(17): 5309-5317. |
| [11] | Baldocchid D, Valentini R, Running S, Oechel W, Dahlman R. Strategies for measuring and modeling carbon dioxide and water vapour fluxes over terrestrial ecosystems. Global Change Biology, 1996, 2(3): 159-168. |
| [12] | LEE X H. On micrometeorological observation of surface-air exchange over tall vegetation. Agriculture and Forest Meteorology, 1998, 91(1/2): 39-49. |
| [13] | Falge E, Baldocchi D, Olson R, Anthoni P, Aubinet M, Bernhofer C, Burba G, Ceulemans R, Dolman H, Granier A, Gross Pa, Grünwald T, Hollinger D, Jensen N O, Katul G, Keronen P, Kowalski A, Ta L C, Law B E, Meyers T, Moncrieff J, Moors E, Munger J W, Pilegaard K, Rannik Ü, Rebmann C, Suyker A, Tenhunen J, Tu K, Verma S, Vesala T, Wilson K, Wofsy S. Gap filling strategies for long term energy flux data sets.Agriculture and Forest Meteorology, 2001, 107(1):71-77. |
| [14] | Li J, Liu Y F, Yang X G, Li J. Studies on water vapor flux characteristic and the relationship with environment factors over a planted coniferous forest in Qianyanzhou station. Acta Ecologica Sinica, 2006, 26(8):2449-2456. |
| [15] | Yu G R, Sun X M. Flux Measurement and Research in Terrestrial Ecosystem in China.Beijing: Science Press, 2008:174-175. |
| [16] | Falge E, Baldocchi D, Olson R, Anthoni P, Aubinet M, Bernhofer C, Burba G, Ceulemans R, Clement R, Dolman H, Granier A, Gross P, Grünwald T, Hollinger D, Jensen N O, Katul G, Keronen P, Kowalski A, Lai C T, Law B E, Meyers T, Moncrieff J, Moors E, Munger J W, Pilegaard K, Rannik Ü, Rebmann C, Suyker A, Tenhunen J, Tu K, Verma S, Vesala T, Wilson K, Wofsy S. Gap filling strategies for defensible annual sums of net ecosystem exchange. Agricultural and Forest Meteorology, 2001, 107(1):43-69. |
| [17] | Huang Z Y, Dong X J, Jiang G M, Yuan W P. Primary studies on the daily dynamic changes of photo-synthesis and transpiration of Salix psammophila. Acta Botannica Boreali-Occidentalia Sinica, 2002, 22 (4): 817-823. |
| [18] | Chapin S F Ⅲ,Matson P A, Mooney H A. Principles of Terrestrial Ecosystem Ecology//Li B, Zhao B, Peng R H, Lu J Z, Jiang L F, Liao C Z, Li C G, Yang X M, Ma T, Wu X W, Ma Z J translations. Beijing: Advanced Education Press, 2005:61-62. |
| [19] | Berbigier P, Bonnefond J M, Mellmann P. CO2 and water vapor fluxes for 2 years above Euro flux forest site. Agricultural and Forest Meteorology, 2001, 108(3): 183-197. |
| [20] | Wang X, Wang C L, Zhou G Y, Yan J H, Sun G, Wang C L. Dry season energy balance of a coniferous and broad-leaved mixed forest at Dinghushan mountain, Southern China. Journal of Tropical and Subtropical Botany, 2005, 13(3):205-210. |
| [21] | Zhao M F, Xiang W H, Tian D L, Zhao Z H,Yan W D,Fang X. The evapotranspiration estimation of planted Chinese fir in Huitong of Hunan Province based on the 3-PG model. Hunan Agricultural Science, 2008(3):158-162. |
| [22] | Wang Z Y. Energy balance and Water Vapor Flux of Snail Control and Schistosomiasis Prevention Forests Ecosystem in Yangtze River Beach Land [D]. Beijing: Chinese Forestry Research, 2008. 70-72. |
| [23] | Yu G R, Wang Q F, Mi N. Ecophysiology of Plant Photosynthesis, Transpiration, and Water Use. Beijing: Science Press, 2010:351-352. |
| [24] | Guo R P, Mo X G. Differences of evapotranspiration on forest, grassland and farmland. Chinese Journal of Applied Ecology, 2007, 18(8):1751-1757. |
| [25] | Yu G R, Niu D, Wang S Q, Wang Q F, Fu Y L, Liu X A, Ren C Y, Mi N, Sun X M,Li Z Q, Li Q K, He H L, Song X, Zhang L M, Zhao F H, Gao L P, Wen X F. Principles of Flux Measurement in Terrestrial Ecosystems. Beijing: Advanced Education Press, 2006:33-34. |
| [26] | Mahrt L, Vickers D. Relationship of area-averaged carbon dioxide and water vapour fluxes to atmospheric variables. Agricultural and Forest Meteorology, 2002, 112(3/4): 195-202. |
| [27] | Jia Zh J, Song Ch Ch, Wang Y S, Huang Y, Shi L Q. Studies on Evapotranspiration over Mire in the Sanjiang Plain. Climatic and Environmental Research, 2007, 12(4):496-502. |
| [28] | Ji X B, Kang E S, Zhao W Z, Chen R S, Jin B W, Zhang Z H. Simulation of the evapotranspiration from irrigational farmlands in the Oases of the Heihe River Basin. Journal of Glacialogy and Geocryology, 2004, 26(6):713-719. |
| [29] | Anthoni P M, Law B E, Unsworth M H. Carbon and water vapor exchange of an open-canopied ponderosa pine ecosystem. Agricultural and Forest Meteorology, 1999, 95(3): 151-168. |
| [30] | Takagi K, Tsuboya T, Takahashi H. Diurnal hystereses of stomatal and bulk surface conductances in relation to vapor pressure deficit in a cool-temperature wetland. Agricultural and Forest Meteorology, 1998, 91(3/4): 177-191. |
| [1] | 温国胜, 张利阳, 张汝民, 王电杰, 张俊. 毛竹光合生理对气候变化的短期响应模拟. 浙江农林大学学报,2011,28(4):555-561. |
| [2] | 张利阳, 温国胜, 王圣杰, 刘兆玲.毛竹光响应模型适用性分析. 浙江农林大学学报,2011,28(2):188-193. |
| [3] | 施建敏, 郭起荣, 杨光耀. 毛竹光合动态研究.林业科学研究,2005,18(5):551-555. |
| [4] | 李雅红, 江洪, 原焕英, 刘源月, 周国模, 余树全. 西天目山毛竹林土壤呼吸特征及其影响因子.生态学报, 2010, 30 (17): 4590-4597. |
| [5] | 肖复明, 范少辉, 汪思龙, 官凤英, 于小军, 申正其. 毛竹、杉木人工林生态系统碳平衡估算. 林业科学, 2010, 46 (11):59-64. |
| [6] | 周国模.毛竹林生态系统中碳储量、固定及其分配与分布的研究 [D]. 浙江:浙江大学,2006. |
| [10] | 刘渡, 李俊, 于强, 同小娟, 欧阳竹.涡度相关观测的能量闭合状况及其对农田蒸散测定的影响.生态学报,2012,32(17):5309-5317. |
| [14] | 李菊, 刘允芬, 杨晓光, 李俊. 千烟洲人工林水汽通量特征及其与环境因子的关系.生态学报,2006,26(8):2449-2456. |
| [15] | 于贵瑞, 孙晓敏. 中国陆地生态系统碳通量观测技术及时空变化特征. 北京:科学出版社,2008:174-175. |
| [17] | 黄振英, 董学军, 蒋高明, 袁文平. 沙柳光合作用和蒸腾作用日动态变化的初步研究. 西北植物学报, 2002, 22(4):817-823. |
| [18] | 蔡平, 马特森, 穆尼著. 陆地生态系统生态学原理//李博, 赵斌, 彭容豪, 陆建忠, 姜丽芬, 廖成章, 杨晓明, 马涛, 吴晓雯, 马志军, 李晨光译.北京:高等教育出版社,2005:61-62. |
| [20] | 王旭, 王春林, 周国逸, 闫俊华, Sun G, 王春林. 鼎湖山针阔混交林旱季能量平衡研究. 热带亚热带植物学报, 2005, 13(3):205-210. |
| [21] | 赵梅芳, 项文化, 田大伦, 赵仲辉, 闫文德, 方晰. 基于3-PG模型的湖南会同杉木人工林蒸发散估算. 湖南农业科学, 2008(3):158-162. |
| [22] | 王昭艳. 长江滩地抑螺防病林生态系统能量平衡与水汽通量研究 [D]. 北京:中国林业科学研究院,2008. 70-72. |
| [23] | 于贵瑞, 王秋凤, 米娜. 植物光合、蒸腾与水分利用的生理生态学.北京: 科学出版社,2010:350-360. |
| [24] | 郭瑞萍, 莫兴国.森林、草地和农田典型植被蒸散量的差异. 应用生态学报,2007:18(8):1751-1757. |
| [25] | 于贵瑞, 牛栋, 王绍强, 王秋凤, 付玉玲, 刘新安, 任传友, 米娜, 孙晓敏, 李正泉, 李庆康, 何洪林, 宋霞, 张雷明, 赵风华, 高鲁鹏, 温学发. 陆地生态系统通量观测的原理与方法.北京:高等教育出版社, 2006:33-34. |
| [27] | 贾志军, 宋长春, 王跃思, 黄耀, 石立庆. 三江平原典型沼泽湿地蒸散量研究.气候与环境研究,2007:12(4):496-502. |
| [28] | 吉喜斌, 康尔泗, 赵文智, 陈仁升, 金博文, 张智慧. 黑河流域山前绿洲灌溉农田蒸散发模拟研究.冰川冻土,2004, 26(6):713-719. |