 2014, Vol. 34
2014, Vol. 34文章信息
- 孙婴婴, 刘立生, 张岁岐
- SUN Yingying, LIU Lisheng, ZHANG Suiqi
- 不同倍性冬小麦种内竞争能力的比较研究
- A comparative study on intraspecific competitiveness of different ploidy wheats
- 生态学报, 2014, 34(16): 4488-4498
- Acta Ecologica Sinica, 2014, 34(16): 4488-4498
- http://dx.doi.org/10.5846/stxb201212301900
-
文章历史
- 收稿日期:2012-12-30
- 网络出版日期:2014-3-19






2. 中国科学院水利部水土保持研究所, 杨凌712100
2. Institute of Soil and Water Conservation, Chinese Academy of Sciences and Ministry of Water Resources Yangling 712100, China
竞争是生物有机体之间的相互作用、塑造植物形态和植物群落结构的主要动力,也是维持自然群落物种多样性、决定生态系统结构和功能的最重要生态过程[1]。Goldberg[2]认为竞争是由植物相互对资源吸收利用的效应而引起的对资源波动的响应和反应,强调资源间接调节的过程。而作物生产是一个种群的过程而非个体的表现[3, 4],单株优势较强的群体中,由于植株间存在强烈的相互竞争,导致个体间存在严重的生物胁迫而使群体表现下降[3, 5, 6]。Donald首先注意到这个问题,提出谷类作物育种的新思路,其核心内容是以群体表现为目标选育理想株型,即在杂草得到有效控制的条件下,高产作物应为弱竞争能力的理想株型,如矮秆、小根系、高收获指数[7, 8, 9]。单一生物种群中竞争能力强的个体比竞争能力弱的个体获得更多的资源,个体大小更大、繁殖后代数更多[10, 11, 12],即种群中强竞争能力个体拥有高适合度。
小麦群体大小与小麦种内竞争有直接关系。在密度很低即群体较小时植株间无明显的竞争作用[3, 13]。Gooding等研究表明小麦产量随着播种量的增加而提高,当播种量增加至一定水平时,产量就开始下降,低播种量可以通过提高分蘖量和穗粒数来补偿群体过小造成的减产[14]。Stephen等指出,随着播种量的增加将产生密集的群体,使某些产量构成要素有所提高,但产量一般不会增长[15]。这些关于如何实现作物种群的高产的研究大多集中在普通小麦之间抗性研究,而对不同密度条件下进化材料群体内个体竞争的研究较少。小麦在从两倍体野生近缘种进化到现代六倍体品种的漫长过程中承受了自然和人工的双重选择压力[16]。从两倍体到四倍体株型从松散状的杂草型向形态紧凑的作物型嬗变,主要是自然选择的结果;从四倍体到六倍体株型从强竞争型向弱竞争型转变,则主要是以产量为目标的人工选择的结果。不同倍体小麦之间种群的大小和竞争能力的强弱对于研究群体与个体协调关系有重要意义。利用3个不同倍性的冬小麦材料,通过不同倍性冬小麦材料在不同水分条件下的单播密度实验,比较株高、生物量、分蘖动态和产量对密度变化的反应,根据各材料对相同群体种内竞争的不同反应,以揭示个体竞争能力、个体适合度与密度之间的内在联系,从小麦进化亲缘关系出发,研究小麦个体的竞争能力和适应能力,为半干旱区小麦高产育种和栽培提供线索,并丰富作物种群生态学的理论研究。
1 实验材料和方法 1.1 试验材料与设计试验选用3种不同染色体倍性的小麦材料,分别是:
染色体组型为AA的栽培一粒(Triticum monococcum,2n=14);
染色体组型为AABB的栽培两粒(Triticum dicoccum,4n=28);
染色体组型为AABBDD的长武134(Triticum aestivum Changwu 134,6n=42)。
试验于2007年10月—2008年6月在陕西省杨凌中国科学院水土保持研究所活动防雨棚内和棚外田间进行。土壤类型为肥熟旱耕人为土,田间最大持水量28%,土壤有机质含量为19.11 g/kg,全N 0.99 g/kg,碱解N 67.00 g/kg,全P 0.87 g/kg,速效P19.40 g/kg,速效K150.40 g/kg。试验田播前一次性施足基肥,施尿素150 kg/hm2(以N计),过磷酸钙120 kg/hm2 (以P2O5计),生育期内不再追肥,田间管理同当地农民习惯。
试验小区面积2 m×3 m,行距20 cm。设两个水分处理,分别为干旱胁迫处理(位于干旱棚内,整个生育期不灌水,也无降水)和正常供水处理(位于棚外田间,1m深度土层含水维持在田间持水量的70%以上);3个密度,分别为低密度(100 万株/hm2),中密度(250 万株/hm2)和高密度(400 万株/hm2)基本苗处理;三因素共18个处理,随机区组排列,3次重复,共计54个小区。
1.2 测定指标及方法 1.2.1 土壤含水量分别于播前及小麦拔节期、抽穗扬花期、收获期,采用CNC503B型土壤中子水分仪每10 cm为一层测定100 cm深度范围内土壤分层含水量,所有分层的土壤含水量加权值即为该小区的土壤容积含水量。
1.2.2 株高、单株分蘖、阶段生物量积累株高采取定株测量的方法,每处理固定10株测定主茎株高,从拔节期开始分别在拔节期、孕穗期、开花期和成熟后进行。植株高度抽穗前以地面至心叶叶尖为准,抽穗后量至穗尖。分蘖采取定点观测的方式,每小区内量取1 m2,确定基本苗数,分别在越冬期和返青期调查分蘖,返青后每隔两周观测一次。在各生育期开始后五天每小区直接拔取五整株,室内去根,在烘箱内105 ℃杀青30 min后 80 ℃烘至恒重,测其生物量累积。
绝对生长速率(AGR,g/d)、相对生长速率(RGR,mg g-1 d-1)由公式 (Altamirano[17];Anten[18])计算得出。
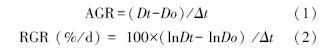
式中,Δt为间隔时间,Dt为某次收割时植物的总干物质量,Do为前一次收割时的植物体干物质量。
1.2.3 计产收获期每小区量取1 m2数其全部穗数,并计算单位面积穗数;将全部地上部分风干后称地上部生物量,将全部籽粒人工搓出风干后称重,用于测定产量。另取10个有代表性主茎穗头,数其穗粒数,籽粒搓下后称穗粒重和千粒重。
1.3 数据整理采用Excel2003进行数据整理;采用SAS-V8处理软件进行统计分析,新复极差法检验处理间的差异显著性水平;sigmaplot10.0绘图。
2 结果分析 2.1 不灌水条件下土壤水分动态变化图 1显示了不灌水条件下不同倍性材料冬小麦主要生育期1 m深度内土壤容积含水量变化,播前土壤水库储存大量水分,3个不同倍性材料的不同密度处理下,土壤容积含水量从播期29%左右降至花期的15%左右,能够保证冬小麦生育期内基本生长要求。

|
| 图 1 干旱条件下不同倍性材料冬小麦主要生育期土壤容积含水量变化 Fig. 1 Dynamic changes of soil water content under drought conditions of different different ploidy materials 2n、4n、6n分别代表两倍体、四倍体和六倍体; D代表不灌水胁迫处理;L、M、H分别代表低密度、中密度和高密度处理 |
由图 2可知,随着倍性增加,冬小麦株高呈现显著下降趋势,且3个材料对于水分的反应类似,即正常供水处理下的株高均显著高于干旱胁迫,且两倍体和四倍体水分间的差异随着生育期的推进而增大,但六倍体在两种水分处理之间差异较小。

|
| 图 2 不同水分密度条件下3个倍性材料株高动态变化 Fig. 2 Dynamic changes of three ploidy materials height under the different water and density conditions D和Ir分别表示干旱和正常供水处理;L、M、H分别代表低密度、中密度和高密度处理;不同大写字母表示相同材料相同水分不同密度间在0.05水平有显著差异,不同小写字母表示相同材料相同密度不同水分间在0.05水平有显著差异 |
不同倍性材料对密度的响应并不一致:两倍体各密度株高从孕穗期开始表现差异,干旱胁迫下高密度平均株高显著大于其他两个密度,正常供水下则以中密度最高;开花期干旱胁迫下株高随密度增加逐渐增高,正常供水下以低密度最高,中密度最低;成熟期无论干旱或是正常供水下,均以中密度最高,分别达到114.55 cm和132.60 cm(图 2)。干旱胁迫下,四倍体株高在拔节期随着密度的增加而增高,而正常供水下则无显著差异;孕穗期两种水分条件下均以高密度最高,但干旱胁迫下低密度和中密度间无显著差异;开花期干旱胁迫下低密度和高密度株高均显著大于中密度,正常供水下各密度处理无显著差异;成熟期干旱胁迫下随着密度增加株高逐渐降低,正常供水下中密度>高密度>低密度,即130.73 cm>127.42 cm>122.98 cm(图 2)。六倍体的株高在干旱胁迫下,从拔节期到开花期之间株高均随着密度的增加逐渐增高,最终各密度株高间无显著差异;而在正常供水下,低密度和中密度在拔节期至开花期株高增加显著,分别从拔节期的次于高密度到开花期的接近和超过高密度株高,成熟期最终株高高密度>中密度>低密度,即87.56 cm> 84.24 cm >75.38 cm(图 2)。
2.3 单株生物量图 3显示三个倍性材料的单株生物量之间并无明显差异。而在相同密度下,单株生物量在越冬期和拔节期不同水分环境里均无显著差异,进入孕穗期以后,均表现为正常供水下单株生物量高于水分胁迫环境下,但随着密度的增加差异逐渐减小。

|
| 图 3 不同水分密度条件下3个倍性材料单株生物量动态变化 Fig. 3 Dynamic changes of three ploidy materials biomass of single plant under the different water and density conditions D和Ir分别表示干旱和正常供水处理;L、M、H分别代表低密度、中密度和高密度处理;不同大写字母表示相同材料相同水分不同密度间在0.05水平有显著差异,不同小写字母表示相同材料相同密度不同水分间在0.05水平有显著差异 |
3 个倍性材料的单株生物量对于密度的反应趋于一致,均表现为随密度的增加逐渐降低。其中干旱胁迫下两倍体栽培一粒孕穗期中密度单株生物量达到2.93 g,显著高于低密度和高密度处理,开花期和成熟期的单株生物量均表现为低密度>中密度>高密度。四倍体不同水分下各时期变化趋势基本一致,越冬期和拔节期无显著差异,孕穗期至成熟期单株生物量均表现为低密度>中密度>高密度。该段生育期内低密度单株生物量增加显著,干旱胁迫和正常供水条件下分别增加了3.65和3.91倍,而高密度处理则增加缓慢,仅分别增加了1.23和1.17倍。六倍体生育期内单株生物量变化趋势和四倍体基本一致。
与单株生物量的变化规律一致,不同倍性的个体生物量AGR和RGR的变化(表 1)也随密度的增加而逐渐降低,差异显著。随着倍性的增加,灌浆期内个体生物量的AGR和RGR逐渐增加,灌浆期生物量的积累主要用于籽粒的形成,也体现了小麦将光合产物向籽粒运转的能力;在水分胁迫和正常供水下不同倍性材料成熟期单株生物量均以四倍体的变异系数最大,分别达到82.72、73.19,以六倍体各密度之间变异最小,分别为61.46、63.51(表 2),说明由于个体竞争和适应能力的不同,三个倍性材料个体生长对不同群体密度的响应存在差异,在相同环境条件下四倍体的个体间竞争能力较强,而六倍体最弱。
| 材料 Material | 处理 Treatments | 绝对生长速率 AGR(Absolute grow rate)/(g/d) | 相对生长速率 RGR(Relative growth rate)/(mg g-1 d-1) | ||||||
| 拔节期 Jointing stage | 孕穗期 Booting stage | 开花期 Flowering stage | 成熟期 Maturity | 拔节期 Jointing stage | 孕穗期 Booting stage | 开花期 Flowering stage | 成熟期 Maturity | ||
| 不同大写字母表示相同材料相同水分不同密度间在 0.05 水平有显著差异 ,不同小写字母表示相同材料相同密度不同水分间在 0.05 水平有显著差异 | |||||||||
| 栽培一粒 | 干旱100 | 0.009Aa | 0.101Bb | 0.223A | 0.149Ab | 1.981Bb | 5.802Bb | 4.26Aa | 1.39Ab |
| Triticum | 干旱250 | 0.007Bb | 0.148Ab | 0.077B | 0.019Bb | 1.732Bb | 7.971Aa | 1.666Ba | 0.316Bb |
| monococcum | 干旱400 | 0.010A | 0.089Bb | 0.029C | 0.009Cb | 2.029Aa | 5.039Bb | 0.908Ba | 0.256Bb |
| 灌水100 | 0.007Bb | 0.358Aa | 0.160A | 0.229Aa | 1.430Ba | 11.293Aa | 1.619Ab | 1.492Aa | |
| 灌水250 | 0.012Aa | 0.167Ba | 0.085B | 0.045Ba | 2.313Aa | 6.703Bb | 1.549Aa | 0.616Ba | |
| 灌水400 | 0.013Aa | 0.136Ba | 0.011C | 0.033Ba | 2.042Aa | 5.778Ba | 0.277Bb | 0.697Ba | |
| 栽培两粒 | 干旱100 | 0.008Bb | 0.174Ab | 0.076Ab | 0.291Ab | 2.095Ab | 8.666Aa | 1.487Ab | 2.885Aa |
| Triticum | 干旱250 | 0.008Bb | 0.120ABb | 0.008Bb | 0.071Ba | 1.621Bb | 6.689ABb | 0.242Bb | 1.576Ba |
| dicoccum | 干旱400 | 0.009ABb | 0.069Bb | 0.004Bb | 0.014Cb | 1.966Ab | 4.599Bb | 0.158Ba | 0.543Ca |
| 灌水100 | 0.013Aa | 0.199Aa | 0.156Aa | 0.348Aa | 2.407Aa | 7.360ABb | 2.253Aa | 2.504Ab | |
| 灌水250 | 0.010Aa | 0.211Aa | 0.051Ba | 0.045Bb | 2.241Aa | 8.489Aa | 0.874Ba | 0.625Bb | |
| 灌水400 | 0.012Aa | 0.158Ba | 0.006Ca | 0.018Ca | 2.432Aa | 6.633Ba | 0.133Cb | 0.378Cb | |
| 长武134 | 干旱100 | 0.005Bb | 0.153Ab | 0.037Ab | 0.250Ab | 0.980Bb | 8.432Ab | 0.876ABa | 3.117Ab |
| Triticum | 干旱250 | 0.006Bb | 0.090Bb | 0.034Aa | 0.083Bb | 1.399ABb | 6.088ABa | 1.153Aa | 1.811Ba |
| aestivum | 干旱400 | 0.010Aa | 0.066Bb | 0.014Ba | 0.033Ca | 1.655Aa | 4.030Bb | 0.561Ba | 1.005Ca |
| Changwu 134 | 灌水100 | 0.008Ba | 0.193Aa | 0.048Aa | 0.406Aa | 1.951ABa | 8.988Aa | 0.913Aa | 3.665Aa |
| 灌水250 | 0.014Aa | 0.137Ba | 0.028Bb | 0.117Ba | 2.318Aa | 5.530Bb | 0.634Bb | 1.793Ba | |
| 灌水400 | 0.010Ba | 0.159ABa | 0.007Cb | 0.031Ca | 1.351Bb | 6.494Ba | 0.155Cb | 0.619Cb | |
| 项目 Items | 栽培一粒 T. monococcum L | 栽培两粒 T. dicoccon Schuebl | 长武134 T. aestivum L Changwu 134 | |||
| 水分胁迫 Drought | 正常供水 Irrigation | 水分胁迫 Drought | 正常供水 Irrigation | 水分胁迫 Drought | 正常供水 Irrigation | |
| 株高Height/% | 5.19 | 1.82 | 6.41 | 3.06 | 0.80 | 7.12 |
| 收获期单株生物量 Biomass of single plant/% | 63.30 | 68.02 | 82.72 | 73.19 | 61.46 | 63.51 |
| 单株分蘖 Tiller number per plant/% | 21.36 | 27.42 | 25.10 | 29.09 | 20.22 | 21.12 |
| 产量Yield/% | 6.73 | 6.01 | 1.98 | 17.12 | 9.07 | 2.46 |
由图 4可以看出在不同水分和密度条件下,各材料变化趋势基本相同,均表现为单峰变化趋势,且低密度>中密度>高密度,正常供水>干旱胁迫。干旱胁迫下,不同群体间单株分蘖数变化缓和,而正常供水下分蘖数量增加变化剧烈,峰值明显。随着密度的增加,三种倍性材料个体分蘖数在减小的同时,最大单株分蘖数出现时间也逐渐提前。两倍体干旱胁迫下低密度最大分蘖数出现最晚(图 4),数量最大。正常供水下低密度和中密度单株分蘖高峰出现较晚,在3月22号左右,分别达到16.3和11.0 个/株,显著高于高密度条件下。四倍体分蘖量明显较两倍体少(图 4)。干旱胁迫下,中密度和高密度下最大分蘖数出现在3月22号左右,分别为6.6和5.2 个/株,低密度则出现在4月5号左右。正常供水下分蘖数明显增加,其中高密度单株分蘖在2月22之前高于低密度和中密度,其分蘖高峰7.5 个/株出现在3月8号左右,而低密度和中密度则为3月22号左右,分别为13.2和8.5 个/株。六倍体长武134在干旱胁迫下单株分蘖增加变化缓和,高密度峰值出现最早(图 4)。正常供水下,低密度单株分蘖高峰12.0 个/株出现在3月22号左右,而中密度和高密度则出现在3月8号左右,分别为10.4和7.8 个/株。以上变化趋势都与两倍体和四倍体相类似。

|
| 图 4 不同水分密度条件下三个倍性材料单株分蘖动态变化 Fig. 4 Dynamic changes of three ploidy materials tiller number per plant under the different water and density conditions D和Ir分别表示干旱和正常供水处理;L、M、H分别代表低密度、中密度和高密度处理;不同大写字母表示相同材料相同水分不同密度间在0.05水平有显著差异,不同小写字母表示相同材料相同密度不同水分间在0.05水平有显著差异 |
在个体生育期分蘖动态变化中,水分胁迫下低密度个体最高分蘖和高密度下最低分蘖均为四倍体栽培两粒,而六倍体长武134表现出相反的趋势。正常供水下,各群体内个体分蘖数以两倍体栽培一粒最高,低密度下以六倍体长武134最低,高密度下则以四倍体栽培两粒最低。不同倍性材料个体竞争能力和对密度的适应能力不同。在相同水分处理下,由不同密度群体间单株分蘖数变异系数可以看出(表 2),四倍体栽培两粒的群体间变异系数最大,说明密度对于该材料的影响最大,而六倍体长武134最小。随着密度的增加,个体间的竞争逐渐增强,密度在抑制个体分蘖的同时,加剧了群体内分蘖的分化,对于竞争能力较强的材料来说,竞争压力随着密度和分蘖的增加逐渐增强,最终导致个体分蘖的剧烈变化。
2.5 产量及产量构成小麦产量由多个因素构成,表 3为3个倍性材料的产量及产量构成因素。其中单位面积穗数不仅受到品种特性的影响,也受到初始群体大小和群体演化过程中内部环境条件的影响。水分胁迫下两倍体和六倍体随着密度的增加,单位面积的穗数也逐渐增加。在中密度和高密度下,两倍体的穗数最多,而四倍体最少;在低密度下,以四倍体穗数最多为472 万穗/hm2,六倍体的穗数最少为433 万穗/hm2。在正常供水下各倍性材料单位面积穗数变化趋势基本相同,均随着密度的增大而增多;而在相同密度下各材料相对变化却不尽相同,低密度下为两倍体>四倍体>六倍体,而在中密度和高密度下则都是两倍体>六倍体>四倍体。
| 材料 Material | 处理 Treatment | 单位面积穗数 /(万/hm2) Spike density | 穗粒数 Kernels per spike | 千粒重/g Thousand kernel weight | 收获指数/% Harvest index | 产量/(kg/hm2) Yield |
| 不同大写字母表示相同材料相同水分不同密度间在 0.05 水平有显著差异 ,不同小写字母表示相同材料相同密度不同水分间在 0.05 水平有显著差异 | ||||||
| 栽培一粒 | 干旱100 | 446Bb | 31.6Ab | 30.230Aa | 28.0Aa | 3693.6Ab |
| Triticum monococcum | 干旱250 | 587Ab | 34.0Aa | 27.665Bb | 27.2Aa | 4218.9Aa |
| 干旱400 | 657Aa | 30.1Aa | 26.004Cb | 25.7Aa | 3896.2Ab | |
| 灌水100 | 601Ba | 35.7Aa | 30.067ABa | 25.1Aa | 4852.2Aa | |
| 灌水250 | 697ABa | 34.0Aa | 32.644Aa | 21.8Aa | 4436.3Aa | |
| 灌水400 | 760Aa | 31.2Aa | 28.654Ba | 23.6Aa | 4984.2Aa | |
| 栽培两粒 | 干旱100 | 472Ab | 32.9Aa | 31.338Aa | 26.2Aa | 4035.2Ab |
| Triticum dicoccum | 干旱250 | 562Aa | 29.8Aba | 27.890Ab | 27.2Aa | 3878.8Ab |
| 干旱400 | 553Ab | 28.3Ba | 27.374Aa | 30.5Aa | 3963.6Ab | |
| 灌水100 | 581Ba | 37.8Aa | 30.400Ba | 28.4Aa | 5750.3Aa | |
| 灌水250 | 630Ba | 32.6Ba | 33.203Aa | 22.5Ab | 4434.7Ba | |
| 灌水400 | 713Aa | 27.9Ca | 29.503Ba | 21.2Ab | 4235.7Ba | |
| 长武134 | 干旱100 | 433Ba | 19.2Ab | 50.181Aa | 34.7Ab | 4339.1Bb |
| Triticum aestivum | 干旱250 | 563Ab | 24.4Aa | 49.057Aa | 32.4Aa | 4895.2Ab |
| Changwu 134 | 干旱400 | 595Aa | 21.1Ab | 48.622Aa | 34.3Aa | 5200.1Ab |
| 灌水100 | 557Ba | 31.5Aa | 49.847Aa | 48.2Aa | 9038.8Aa | |
| 灌水250 | 691ABa | 26.9Ba | 50.406Aa | 41.6Aa | 8858.4Aa | |
| 灌水400 | 723Aa | 30.4ABa | 48.329Aa | 39.3Aa | 8605.9Aa | |
水分胁迫下,四倍体随着密度的增加穗粒数逐渐减少,而其他两个倍性材料则均是中密度下穗粒数最多;正常供水下,栽培一粒和栽培两粒穗粒数均随着密度的增加逐渐减少,六倍体长武134则表现为低密度>高密度>中密度,即31.5>30.4>26.9。在相同密度下,除四倍体的高密度处理外,3个倍性材料的低、中密度在正常供水下的穗粒数较水分胁迫下均有所增加。两种水分条件下,现代品种长武134的穗粒数均低于两原始品种。
无论在干旱胁迫还是正常供水下,千粒重随着染色体倍数从2n→6n的增加逐渐增加;水分胁迫下三个倍性材料千粒重均随密度的增加而降低;正常供水处理各材料均以中密度下的千粒重最高,分别达到32.64、33.2 g和50.41 g;同时正常供水下除六倍体的高密度处理外,各倍性材料的中密度和高密度处理的千粒重均高于对应密度下水分胁迫处理。
收获指数反映了植物后期光合产物的积累和分配状况。水分胁迫下两倍体的收获指数随着密度的增加减小,而四倍体的趋势完全相反,六倍体现代品种则以低密度下最高为34.7%,中密度的收获指数最低为34.3%,差异并不明显。正常供水下四倍体和六倍体收获指数都随着密度的增加而减小,而两倍体则以低密度的最高,中密度最低。两种水分条件下收获指数都随着染色体倍数的增加逐渐提高。
水分胁迫处理下,两倍体和四倍体分别以中密度和低密度下的产量最高,六倍体的产量则随着密度的增加而增加。三个倍性材料水分胁迫下不同密度的产量变异系数分别为6.73%、1.98%和9.07%,说明六倍体受密度的影响最为明显。正常供水下,四倍体和六倍体的产量随着密度的增加而减少,而两倍体则以高密度下产量最高。三个倍性材料中,四倍体产量变异系数达到17.12%,六倍体的产量变异系数仅为2.46%。正常供水各处理的产量均高于水分胁迫处理,尤其在低密度下,随着染色体倍性的增加产量分别增加了31.37%、42.50%和108.31%。两种水分条件下均以六倍体的产量最高,两倍体和四倍体差异不显著。
3 讨论 3.1 水分和密度对不同倍性小麦个体生长的影响个体的生长发育能直接体现群体内竞争状况[2]。在本研究中,种群密度的增加使得小麦生长所需的资源缺乏不断放大,个体间的竞争更加激烈,生物量与分蘖受到明显的限制,增长空间减小,生物量累积速率下降。水分亏缺下群体建立的延迟,使得个体分蘖数峰值出现时间偏后。充足的水分促进个体各构件的生长,群体间差异也随之显著增加,同时个体间的竞争也倾向于对光照的竞争,最直接的表现为株高的变化,收获期两倍体和四倍体均表现为中密度下株高最高,而六倍体则表现为高密度下最高。梁志刚等认为:小麦播种密度在225—450 万株/hm2范围内,随着播种密度的增加株高增加以实现有限资源下对光照的利用[19],这一结论与本文六倍体材料上的研究结果一致,而原始品种因为其高的个体竞争力和低的群体耐密性,其株高对密度的响应略有不同。多个研究均表明在一定范围内单位面积穗数随密度增大而增加,干物质的积累也呈上升趋势,但当密度增大到一定程度后干物质积累不增反降[20, 21],这些均与本文的研究结果一致。
3.2 不同倍性小麦产量对于水分和密度的响应小麦生育期内的竞争结果最终反映在群体产量上:本研究中随着群体增大,竞争加强,个体繁殖力逐渐减小。虽然群体增大单位面积穗数显著增加,但是由于对资源的竞争增强,个体竞争的加强,使得花后干物质累积受限,最终导致个体生殖能力下降,穗粒数和千粒重显著降低,最后导致产量并非随着群体的增大而增加,且水分亏缺下表现尤为明显。这些结果表明并产量与群体大小关系并非完全成正向线性关系。刘丽平等人的研究认为密度增大总茎数和干物质积累量显著增加,籽粒产量则呈低-高-低的变化趋势[22]。研究表明,在穗数随着种植密度的增加而增加的同时,小麦的穗粒数和千粒重下降[23, 24, 25];但马东钦等[26]认为,种植密度对穗数和穗粒数影响的显著程度非常高,对千粒重的影响不显著,与本研究得到的结果并不一致。正常供水下四倍体和六倍体均以低密度即小群体下的产量最高,证明适宜的密度条件下通过水分的合理运筹,提高生育中后期群体光合生产能力,增加花后干物质积累量及其所占比重在节水的前提下同样可以获得高产量。
3.3 小麦在倍体演化进程中竞争能力与群体生产力的变化本研究中,长武134的分蘖数和株高明显低于两倍体和四倍体,个体竞争资源的能力下降,花后生物量累积速率上则具有明显优势,而四倍体栽培两粒则表现出最强烈的个体竞争能力。两种水分条件下均以长武134群体产量最高,栽培一粒和栽培两粒两个较原始品种间无显著差异。长武134作为人工培育出的对人工栽培环境高度适合的小麦品种,虽然穗粒数和穗数没有明显优势,但是千粒重达到50 g左右,收获指数接近50%,对于生殖生长的投入比例增加,在维持生物量的基础上提高了产量。现代品种长武134在干旱条件下各密度处理的产量下降最显著,干旱胁迫下对密度的反应最强烈,说明干旱对其影响最大,即其抗旱能力最低,这与黄明丽、李秧秧等人的研究结果相一致[27, 28]。许多研究表明,小麦在倍体演化进程中,个体竞争能力的改变与群体生产能力息息相关,现代品种正因其低的个体竞争能力和高群体协调能力才更适宜农业栽培[28, 29]。本文的结果也说明了这一结论。
在资源有限条件下,强竞争能力的获得是以牺牲籽实产量为代价的,竞争能力越强,这种代价就越高。因此,作物品种竞争能力与生产能力之间形成了一种负相关关系。现代小麦品种的选育成功,是因为减少了为获得竞争能力而产生的生长冗余,使更多的资源释放出来用于产量形成,虽然其个体生殖竞争能力并非突出,但是在繁殖能力上的投入最高。六倍体长武134作为长期人工选择的结果,属于培育出的竞争能力小,繁殖能力高的品种,能够通过减弱投向竞争能力方向的资源而获得更高产量,更接近于理想株型群体。
个体与群体的关系目前仍是小麦育种的关键问题,如何在群体水平上实现产量最大化仍是重要课题[5, 10, 30, 31, 32],在资源有限的条件下更为突出。水资源的短缺是限制旱地农业发展的重要因素,如何维持水资源在群体和个体间的分配,是提高籽粒产量和水分利用效率的关键所在。合理控制生产种群的大小和结构,减小个体为获得竞争能力而向营养器官诸如根、茎、叶及分蘖等部位分配的比例,从而增大资源向繁殖部位增加的比例,以实现有限水资源的集约高效利用和群体上的高产,应成为小麦育种的一项重要育种目标。
| [1] | Li B, Chen J K, Watkinson A R.A literature review on plant competition. Chinese Bulletin of Bobtany, 1998, 15(4):18-29. |
| [2] | Goldberg D E. Components of resource competition in plant communities//Gacre J B, Tilman D, eds,Perspectives on Plant Competition. San Diego: Academic Press, 1990: 27-49. |
| [3] | Zhang R, Zhang D Y, Yuan B Z, Lin K,Wei H. A study of the relationship between competitive ability and productive performance of spring wheat in semi arid regions of Loess Plateau. Acta Phytoecologica Sinica, 1999, 23(3): 205-210. |
| [4] | Zhao S L, Li F M, Zhang D Y, Duan S S.Crop production is a population process.Acta Ecologica Sinica, 1997, 17(1): 100-104. |
| [5] | Zhang D Y, Sun G J, Jiang X H. Donald's ideotype and growth redundancy: a game theoretical analysis.Field Crops Researsch,1999,61(2): 179-187. |
| [6] | Li F M, Yan X, Guo A H, Shan L.A discussion on the non-hydraulic root-sourced signals and life history strategy of wheat crops. Acta Ecologica Sinica, 2000, 20(3): 510-513. |
| [7] | Donald C M. The breeding of crop ideotypes. Euphytica,1968,17(3):385-403. |
| [8] | Donald C M. Competitive plants, communal plants, and yield in wheat crops//Evans L T, Peacock W J, eds. Wheat Science-Today and Tomorrow. Cambridge: Cambridge University Press, 1981: 223-247. |
| [9] | Donald C M, Hamblin J. The convergent evolution of annual seed crops in agriculture. Advances in Agronomy, 1983, 36:97-143. |
| [10] | Weiner J, Solbrig O T. The meaning and measurement of size hierarchies in plant populations. Oecologia, 1984. 61(3): 334-336. |
| [11] | Rhainds M, Gries G, Min M M. Size- and density-dependent reproductive success of bagworms, Metisa plana. Entomologia Experi mentalis et Applicata, 1999, 91(3): 375-383. |
| [12] | Zhang R, Sun G J,Li F M. Definition of redundancy and the ecological mechanism of its occurrence.Acta Botanica Boreali-Occidentalia Sinca, 2003, 23(5): 844-851. |
| [13] | Li Z F, Wang G. The effects of intraspecific competition on the relationship between height growth and biomass of annual plants. Journal of Lanzhou University: Natural Sciences, 2002, 38(2): 141-145. |
| [14] | Gooding M J, Pinyosinwat A, Ellis R H. Responses of wheat grain yield and quality to seed rate. The Journal of Agriculture Science, 2002, 138(3): 317-331. |
| [15] | Stephen R C, Saville D J, Drewitt E G. Effects of wheat seed rate and fertilizer nitrogen application practices on populations, grain yield components and grain yields of wheat (Triticum aestivum). New Zealand Journal of Crop Horticultural Science, 2005, 33(2): 125-138. |
| [16] | Ken-ichi Tanno, Willcox G. How fast was wild wheat domesticated?. Science, 2006, 311(31): 1886-1886. |
| [17] | Altamirano M, Flores-Moya A, Figueroa F L. Effects of UV radiation and temperature on growth of germlings of three species of Fucus (Phaeophyceae). Aquatic Botany, 2003, 75(1): 9-20. |
| [18] | Anten N P R, Ackelrly D D. A new method of growth analysis for plants that experience periodic losses of leaf mass. Functional Ecology, 2001, 15(6): 804-811. |
| [19] | Liang Z G, Wang J L, Cui H H, Wang Y Z, Jing H. Influence of high temperature before winter and sowing time and density on dynamic growth of individual and population at seedling stage of wheat. Chinese Agricultural Science Bulletin, 2007, 23(8): 185-189 |
| [20] | Guo T C, Cha F N, Ma D Y,Song X,Yue Y J.Effects of plant density on the accumulation and transfer of dry matter and nitrogen and grain yield of two winter wheat cultivars with different spike types. Acta Agriculturae Boreali-Sinica, 2007, 22(6): 152-156. |
| [21] | Cong X J, Wu K, Qian Z G, Sun X Y, Sha Y, Wang C.Effects of plant densities on accumulation of dry matter and population trends of Taishan 21 under the high yield condition.Shandong Agricultural Science, 2004(4): 16-18. |
| [22] | Liu L P, Ouyang Z, Wu L F, Sun Z Z. Effect of irrigation schedules on population quality and grain yield of winter wheat under different densities. Journal of Triticeae Crops, 2011, 31(6): 1116-1122. |
| [23] | Zhu C L, Li R N, Zhang B J, Zhang ZM, Zhao Y P, Wang Y L, Han S M. Effect of planting density on growth characters and yield of winter wheat variety of big spike in dryland. Acta Agriculturae Boreali-Occidentalis Sinica, 2010, 19(1): 71-75. |
| [24] | Wen H X, Feng W S, Duan G H,Yu S P, Zhang X P. Effects of different planting densities on grain-filling characteristics and yield of winter wheat. Acta Agricuhurae Jiangxi, 2009, 21(12): 23-25. |
| [25] | Tian W Z, Wen H X, Gao H T, Yang H Q, Yu S P, Duan G H, Zhang S L. Effects of sowing date, density and their combination on yield of wheat[J]. Journal of Henan Agricultural Sciences, 2011, 40(2): 45-49. |
| [26] | Ma D Q, Wang X W, Zhu Y P, Xu L J, Ma C Y, Sun W X, Guo C Y, Zhan K H. Effect of sowing time and seedling density on the yield formation of a new wheat cultivar Yunong 202. Chinese Agricultural Science Bulletin, 2010, 26(1): 91-94. |
| [27] | Li Y Y, Shao M A. Physi-ecoological response of spring wheat root to water and nitrogen. Plant Nutrition and Fertilizer Science, 2000, 6(4): 383-388. |
| [28] | Huang M L, Deng X P, Zhou S L, Zhao Y Z. Grain yield and water use efficiency of diploid, tetraploid and hexaploid wheats. Acta Ecologica Sinica, 2011, 27(3): 1113-1121. |
| [29] | Austin R B, Morgan C L, Ford M A, Bhagwat S G. Flag leaf photosynthesis of Triticum aestivum and related diploid and tetraploid species. Annals Botany, 1982, 49(2): 177-189. |
| [30] | Reif J C, Zhang P, Dreisigacker S, Warburton M L, van Ginkel M, Hoisington D, Bohn M, Melchinger A E. Wheat genetic diversity trends during domestication and breeding. Theoretical and Applied Genetics, 2005, 110(5): 859-864. |
| [31] | Li P F, Cheng Z G, Zhao H, Zhang X F, Li J N, Wang S M, Xiong Y C. Current progress in plant ideotype research of dryland wheat (Triticum aestivum L.). Acta Ecologica Sinica, 2011, 31(9): 2631-2640. |
| [32] | Wu Y C, Zhou S L, Zhang Y P, Wang Z M. Discussion of plant shapes of high-yield and water-saving wheat. Agriculture Research in the Arid Areas, 2005, 23(2): 126-129. |
| [1] | 李博, 陈家宽, 沃金森 A·R. 植物竞争研究进展. 植物学通报, 1998, 15(4): 18-29. |
| [3] | 张荣, 张大勇, 原保忠, 林魁, 魏虹. 半干旱区春小麦品种竞争能力与产量关系的研究.植物生态学报, 1999, 23(3): 205-210. |
| [4] | 赵松岭, 李凤民, 张大勇, 段舜山. 作物生产是一个种群过程. 生态学报, 1997,17(1): 100-104. |
| [6] | 李凤民, 鄢珣, 郭安红, 山仑. 试论麦类作物非水力根信号与生活史对策. 生态学报, 2000,20(3):510-513. |
| [12] | 张荣, 孙国钧, 李凤民. 冗余概念的界定与冗余产生的生态学机制. 西北植物学报, 2003,23(5):844-851. |
| [13] | 李仲芳, 王刚. 种内竞争对一年生植物高生长与生物量关系的影响. 兰州大学学报: 自然科学版, 2002, 38(2): 141-146. |
| [19] | 梁志刚, 王娟玲, 崔欢虎, 王裕智, 靖华. 冬前高温和播期密度对小麦苗期个体及群体生长的影响. 中国农学通报, 2007, 23(8): 185-189. |
| [20] | 郭天财, 查菲娜, 马冬云, 宋晓, 岳艳军. 种植密度对两种穗型冬小麦品种干物质和氮素积累运转及产量的影响.华北农学报, 2007, 22(6): 152-156. |
| [21] | 丛新军, 吴科, 钱兆国, 孙宪印, 沙英, 王超. 超高产条件下种植密度对泰山1号群体动态干物质积累和产量的影响. 山东农业科学, 2004, (4): 16-18. |
| [22] | 刘丽平, 欧阳竹, 武兰芳, 孙振中. 灌溉模式对不同密度小麦群体质量和产量的影响. 麦类作物学报, 2011, 31(6): 1116-1122. |
| [23] | 朱翠林, 李锐宁, 张保军, 张正茂, 赵永萍, 王玉玲, 韩思明. 旱肥地密度对大穗型冬小麦品种生育特性及产量的影响. 西北农业学报, 2010, 19(1): 71-75. |
| [24] | 温红霞, 冯伟森, 段国辉, 余四平, 张学品. 不同播种密度对冬小麦灌浆特性及产量的影响. 江西农业学报, 2009, 21(12): 23-25. |
| [25] | 田文仲, 温红霞, 高海涛, 杨洪强, 余四平, 段国辉, 张少澜. 不同播期、播种密度及其互作对小麦产量的影响. 河南农业科学, 2011, 40(2): 45-49. |
| [26] | 马东钦, 王晓伟, 朱有朋, 许兰杰, 马彩艳, 孙文鑫, 郭春燕, 詹克慧. 播种期和种植密度对小麦新品种豫农202 产量构成的影响. 中国农学通报, 2010, 26(1): 91-94. |
| [27] | 李秧秧, 邵明安. 小麦根系对水分和氮肥的生理生态反应. 植物营养与肥料学报, 2000, 6(4): 383-388. |
| [28] | 黄明丽, 邓西平, 周生路, 赵玉宗. 二倍体、四倍体和六倍体小麦产量及水分利用效率. 生态学报, 2011,27(3): 1113-1120. |
| [31] | 李朴芳, 程正国, 赵鸿, 张小丰, 李冀南, 王绍明, 熊友才. 旱地小麦理想株型研究进展. 生态学报, 2011, 31(9): 2631-2640. |
| [32] | 吴永成, 周顺利, 张永平, 王志敏. 节水高产小麦理想全株型探讨. 干旱地区农业研究, 2005, 23(2): 126-129. |