 2014, Vol. 34
2014, Vol. 34文章信息
- 姜丽娜, 张黛静, 宋飞, 刘佩, 樊婷婷, 余海波, 李春喜
- JIANG Lina, ZHANG Daijing, SONG Fei, LIU Pei, FAN Tingting, YU Haibo, LI Chunxi
- 不同品种小麦叶片对拔节期低温的生理响应及抗寒性评价
- Evaluation of cold resistance of different wheat varieties based on physiological responses of leaves to low temperature at the jointing stage
- 生态学报, 2014, 34(15): 4251-4261
- Acta Ecologica Sinica, 2014, 34(15): 4251-4261
- http://dx.doi.org/10.5846/stxb201307271962
-
文章历史
- 收稿日期:2013-7-27
- 修订日期:2014-3-8






低温冷(冻)害是黄淮海麦区频发的自然灾害之一,严重影响并制约着小麦的生长发育及产量的形成[1, 2]。低温胁迫后,植物体通过形态结构、内含物质和光合作用等一系列复杂的生理生化变化,对低温环境进行响应[3, 4, 5, 6]。研究表明,植物的抗寒性与细胞膜的流动性[7]、膜脂过氧化程度[8, 9]、抗氧化酶活性[9, 10, 11]、渗透调节物质含量[12, 13]密切相关。植物抗寒性是多基因控制的数量性状,单一的生理指标难以全面准确反映抗寒性的强弱,可结合与抗寒性有关的多个指标,利用隶属函数法[14, 15]、加权法[16]、模糊评判法[17]、极点排序法[18]等进行抗寒性评价。其中,加权法和模糊评判法需要人为确定指标的权重值,而权重值的大小直接影响评判的结果。隶属函数法是根据模糊数学原理,计算的隶属函数值是多项指标综合的结果,能比较准确地反映出品种间的抗寒性差异,在小麦[14]、果桑[15]、甘蔗[19] 、葡萄[20]等均有应用。极点排序法,是将与抗寒性相关的多个指标进行权重分配,通过矩阵复合运算获得综合评价结果,已在甘蔗[18]、辣椒[21]、黄瓜[22]等进行了应用,而在小麦抗寒上未见报道。本文以黄淮海麦区参加区试的24个小麦品种为材料,在人工气候室模拟拔节期低温,分析其对叶片生理的影响。以低温胁迫后各性状的相对值作为抗寒性评价指标[14, 23],通过隶属函数法和极点排序法对参试品种抗寒性进行鉴定,以筛选应对春季低温胁迫的抗寒性品种及抗寒性鉴定指标,为增强黄淮海区域小麦减灾抗灾能力提供理论支持。
1 材料与方法 1.1 供试材料与试验设计供试小麦品种及其供种单位如表 1所示。
| 编号 No. | 品种名称 Variety | 供种单位 Variety source | 编号 No. | 品种名称 Variety | 供种单位 Variety source |
| 1 | 石麦19 | 河北省石家庄市农业科学研究院 | 13 | 山农055843 | 山东农业大学 |
| 2 | 石优20 | 河北省石家庄市农林科学研究所 | 14 | 良星99 | 山东省德州市良星种子研究所 |
| 3 | 石H083-366 | 河北省石家庄市农业科学研究院 | 15 | 良星619 | 山东省德州市良星种子研究所 |
| 4 | B05-6507 | 河北省石家庄市农业科学研究院 | 16 | 尧麦16 | 山西省农业科学院 |
| 5 | B07-4056 | 河北省石家庄市农业科学研究院 | 17 | 舜麦1718 | 山西省农业科学院 |
| 6 | 石06-6136 | 河北省石家庄市农业科学研究院 | 18 | 陕农509 | 西北农林科技大学 |
| 7 | 石B05-7388 | 河北省石家庄市农业科学研究院 | 19 | 徐麦4036 | 徐州农业科学研究 |
| 8 | 石4185 | 河北省石家庄市农业科学研究院 | 20 | 丰德存麦1号 | 河南省天存小麦改良技术研究所 |
| 9 | 冀麦585 | 河北省农林科学院 | 21 | 宿553 | 宿州市农业科学院 |
| 10 | 偃展4110 | 河南省豫西农作物品种展览中心 | 22 | A-9 | 河北省农林科学院 |
| 11 | 中原6号 | 河南谷得科技种业有限公司 | 23 | B-33 | 河北省农林科学院 |
| 12 | 山农05-066 | 山东农业大学 | 24 | C-44 | 河北省农林科学院 |
2 010年10月24日在河南师范大学网室进行盆栽种植。各品种选取完整无损、大小均匀的小麦籽粒,播入米氏盆(h19cm×φ23cm)中,每盆装土2.5kg,播种30粒。随机排列,每品种4盆。三叶期每盆定苗15株,常规管理。于2011年4月5日将低温组(每品种2盆)移入人工气候室中,16℃昼(16h,12000lx)/ 10℃夜条件下正常生长1d,然后在4℃昼(16h,12000lx)/ 4℃夜低温条件下连续处理3d。对照组(每品种2盆)仍在网室中正常生长,环境温度如图 1所示。处理结束后,分别取低温和对照各品种小麦的倒一叶用于生理指标的测定,每品种重复2次。

|
| 图 1 对照组环境温度 Fig. 1 The environment temperature of Control |
相对电导率采用电导率仪法[24],可溶性糖含量采用蒽酮比色法[24],可溶性蛋白含量采用考马斯亮蓝G-250比色法[24],游离脯氨酸含量采用磺基水杨酸浸提-酸性茚三酮显色法[25],丙二醛(MDA)含量采用TBA比色法[26],超氧化物歧化酶(SOD)活性采用NBT比色法[24],过氧化物酶(POD)活性采用愈创木酚比色法[27]。
1.3 数据分析 1.3.1 性状相对值以公式1[15]计算性状相对值xj作为评价品种抗寒性的指标:
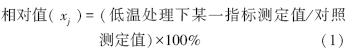
以公式2和公式3计算各品种相对性状的隶属度值[14, 15]。其中,与抗寒性呈正相关的各指标(可溶性糖含量、可溶性蛋白含量、游离脯氨酸含量及SOD活性、POD活性)隶属度值以公式2计算,与抗寒性呈负相关的各指标(MDA含量和相对电导率)隶属度值以公式3计算。
每一品种相对性状隶属度值的平均数即为该品种的平均隶属度值:
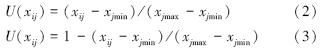
式中,U(xij)为i品种j指标的隶属函数值,xij为i品种j指标的相对值,xjmin和xjmax为各品种j指标相对值的最小值和最大值。
1.3.3 综合排序值(1) 将指标相对性状值划分为5级,每一指标的得分级差(每得1分之差)记为D,以公式4计算各相对指标的得分级差Dj[18]。
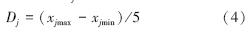
(2) 对每一品种所测指标相对值进行分级,使每一品种的每一指标都得到相应级别值Eij。当指标与抗寒性呈正相关时,该指标的Eij值以公式5计算;当指标与抗寒性呈负相关时,该指标的Eij值以公式6计算:
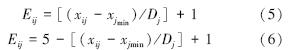
(3) 指标权重系数Bj为每一指标在综合评价中的权重,以公式7进行计算:

(4) 每一品种综合排序值Vi以公式8进行计算:
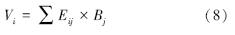
利用Microsoft Excel 2003和SPSS 13.0进行数据处理和作图。
2 结果与分析 2.1 拔节期低温对小麦叶片生理指标的影响 2.1.1 相对电导率正常生长条件下,参试的24个小麦品种叶片相对电导率平均为(23.08±3.99)%(图 2),品种间差异极显著(P<0.01),以山农05-066(12号)叶片相对电导率表现最低,石麦19(1号)最高。低温处理后,叶片电解质外渗增加,相对电导率极显著高于对照(P<0.01)。低温处理后,不同品种小麦叶片相对电导率的极差值增加,变异系数没有明显变化。与对照相比,在供试的24个品种中,山农05-066(12号)叶片相对电导率增幅最高(291.68%),而石麦19(1号)增幅最低,仅为72.63%。

|
| 图 2 低温胁迫下不同小麦品种叶片相对电导率 Fig. 2 Relative electrical conductivity in wheat leaves in response to low temperature stress |
对照组叶片可溶性糖含量为(18.05±4.41) mg/g干重(图 3),品种间差异极显著(P<0.01)。低温促进了小麦叶片可溶性糖的积累,低温处理后小麦叶片可溶性糖含量平均为(25.40±5.89) mg/g干重,极显著高于对照(P<0.01),且不同品种间极差值增加,而变异系数没有明显变化。与对照相比,良星619(15号)叶片可溶性糖含量增加了97.80%,而石06-6136(6号)仅增加了4%。

|
| 图 3 低温胁迫下不同小麦品种叶片可溶性糖含量 Fig. 3 Soluble sugar content in wheat leaves in response to low temperature stress |
对照组小麦叶片可溶性蛋白含量平均为(6.09±1.98) mg/g鲜重(图 4),品种间差异极显著(P<0.01)。低温处理后叶片可溶性蛋白含量为(2.65±1.35) mg/g鲜重,极显著低于对照(P<0.01),且品种间可溶性蛋白含量极差值亦低于对照,而品种间变异系数高于对照。与对照相比,石4185(8号)叶片可溶性蛋白含量降低幅度最高(86.73%),而山农05-066(12号)仅降低了5.58%。

|
| 图 4 低温胁迫下不同小麦品种叶片可溶性蛋白含量 Fig. 4 Soluble protein content in wheat leaves in response to low temperature stress |
对照组小麦叶片游离脯氨酸含量平均为(131.36±73.70) μg/g鲜重(图 5),品种间差异极显著(P<0.01)。低温处理后,叶片游离脯氨酸含量增加,平均为(236.33±96.19) μg/g鲜重,极显著高于对照(P<0.01),其品种间极差值亦高于对照,而变异系数低于对照。与对照相比,良星619(15号)叶片脯氨酸含量增加了350.43%,石4185(8号)仅增加了0.39%。

|
| 图 5 低温胁迫下不同小麦品种叶片游离脯氨酸含量 Fig. 5 Free proline content in wheat leaves in response to low temperature stress |
对照组小麦叶片MDA含量平均为(1.06±0.29) μmol/g鲜重(图 6),品种间差异极显著(P<0.01)。低温处理后,叶片MDA含量平均值为(1.68±0.46) μmol/g鲜重,极显著高于对照(P<0.01),且品种间极差值高于对照,而变异系数无明显变化。与对照相比,石优20(2号)叶片MDA含量增加了318.32%,而石H083-363(3号)仅增加了10.34%。

|
| 图 6 低温胁迫下不同小麦品种叶片MDA含量 Fig. 6 MDA content in wheat leaves in response to low temperature stress |
对照组叶片SOD活性平均为(278.27±70.61) U/g鲜重(图 7),品种间差异极显著(P<0.01)。低温处理后,叶片SOD活性为(349.75±45.96) U/g鲜重,极显著高于对照(P<0.01),且品种间极差值和变异系数均低于对照。与对照相比,石H083-363(3号)叶片SOD活性增加了171.85%,而徐麦4036(19号)仅增加了1.74%。

|
| 图 7 低温胁迫下不同小麦品种叶片SOD活性 Fig. 7 SOD activity in wheat leaves in response to low temperature stress |
对照组叶片POD活性平均为(59.57±22.83) U/g鲜重(图 8),品种间差异极显著(P<0.01)。低温 处理后,叶片POD活性为(103.50±40.88) U/g鲜重,极显著高于对照(P<0.01),且品种间极差值高于对照,但变异系数未有明显变化。与对照相比,B07-4056(5号)增加了388.29%,而山农05-066(12号)仅增加了10.00%。

|
| 图 8 低温胁迫下不同小麦品种叶片POD活性 Fig. 8 POD activity in wheat leaves in response to low temperature stress |
计算各指标的相对性状值列于表 2。低温胁迫后,相对电导率的相对值和相对脯氨酸含量较高,平均值达到253.27%和223.47%,其次是相对POD活性和相对MDA含量,而相对可溶性蛋白含量较低。从各性状相对值的变异系数来看,相对脯氨酸含量、相对可溶性蛋白含量、相对POD活性的变异系数较高,均高于50%,其次是相对MDA含量和相对SOD活性。
| 编号 No. | 品种名称Variety | x1 | x2 | x3 | x4 | x5 | x6 | x7 |
| x1、x2、x3、x4、x5、x6和x7分别表示相对电导率、可溶性糖含量、可溶性蛋白含量、脯氨酸含量、MDA含量、SOD活性和POD活性的相对值 | ||||||||
| 1 | 石麦19 | 172.63 | 118.05 | 54.55 | 192.39 | 141.94 | 156.50 | 129.92 |
| 2 | 石优20 | 252.86 | 136.93 | 23.13 | 122.00 | 418.33 | 119.18 | 138.67 |
| 3 | 石H083-366 | 209.16 | 150.20 | 25.13 | 323.35 | 110.33 | 271.85 | 155.56 |
| 4 | B05-6507 | 193.81 | 107.40 | 28.38 | 139.06 | 168.06 | 113.37 | 130.00 |
| 5 | B07-4056 | 207.55 | 143.03 | 54.40 | 162.71 | 123.38 | 112.55 | 488.89 |
| 6 | 石06-6136 | 246.86 | 104.00 | 25.56 | 120.70 | 268.85 | 104.42 | 125.00 |
| 7 | 石B05-7388 | 293.27 | 143.88 | 29.44 | 120.86 | 115.05 | 117.00 | 210.00 |
| 8 | 石4185 | 314.51 | 132.62 | 13.27 | 100.39 | 124.22 | 111.44 | 133.93 |
| 9 | 冀麦585 | 302.34 | 147.79 | 54.19 | 116.18 | 114.80 | 107.75 | 187.50 |
| 10 | 偃展4110 | 203.78 | 128.98 | 24.10 | 120.50 | 123.22 | 132.69 | 126.62 |
| 11 | 中原6号 | 234.85 | 177.75 | 48.19 | 121.25 | 130.83 | 271.10 | 120.63 |
| 12 | 山农05-066 | 391.68 | 120.11 | 94.42 | 128.79 | 165.25 | 153.32 | 110.00 |
| 13 | 山农055843 | 324.48 | 189.80 | 94.14 | 334.69 | 187.02 | 116.15 | 153.33 |
| 14 | 良星99 | 225.44 | 152.12 | 34.59 | 436.92 | 139.65 | 157.79 | 150.00 |
| 15 | 良星619 | 248.09 | 197.80 | 55.23 | 450.43 | 134.69 | 111.34 | 208.33 |
| 16 | 尧麦16 | 223.71 | 126.21 | 19.86 | 313.24 | 127.30 | 107.67 | 186.11 |
| 17 | 舜麦1718 | 309.04 | 171.43 | 57.26 | 377.56 | 299.73 | 117.72 | 138.89 |
| 18 | 陕农509 | 255.66 | 195.60 | 90.48 | 137.07 | 188.22 | 126.24 | 177.48 |
| 19 | 徐麦4036 | 231.33 | 113.00 | 35.53 | 302.73 | 182.64 | 101.74 | 297.00 |
| 20 | 丰德存麦1号 | 288.15 | 129.81 | 25.82 | 446.15 | 133.00 | 132.66 | 440.00 |
| 21 | 宿553 | 227.39 | 144.65 | 38.70 | 362.08 | 158.97 | 108.31 | 277.55 |
| 22 | A-9 | 273.77 | 164.25 | 86.31 | 165.69 | 128.08 | 117.71 | 165.28 |
| 23 | B-33 | 211.46 | 127.03 | 39.50 | 153.60 | 261.67 | 123.77 | 125.00 |
| 24 | C-44 | 236.59 | 115.42 | 69.17 | 115.03 | 116.72 | 116.46 | 157.14 |
| 平均 Average | 253.27 | 143.24 | 46.72 | 223.47 | 169.25 | 133.70 | 188.87 | |
| 标准差 Standard deviation | 50.43 | 27.38 | 24.81 | 123.99 | 74.00 | 45.18 | 97.12 | |
| 变异系数 Coefficient of variation | 19.91 | 19.11 | 53.09 | 55.48 | 43.72 | 33.79 | 51.42 | |
根据相对性状值,计算参试品种的平均隶属度值为0.19—0.63,综合排序值为1.66—4.08(图 9)。平均隶属度值和综合排序值高,该品种的抗寒性强;反之,该品种的抗寒性弱。根据平均隶属度值和综合排序值对参试品种进行排序,结果不完全相同,二者均以良星619表现最高,石优20表现最低。

|
| 图 9 各品种的平均隶属度值和综合排序值及其聚类类别 Fig. 9 Average of subordinate function and comprehensive score of each variety and cluster membership |
以平均隶属度值和综合排序值对参试品种进行K-Means聚类,经过4次聚类迭代后将参试品种聚为5类,各类中心值列于表 3。第Ⅰ类综合排序值和平均隶属度值均较高,抗低温能力最强;第Ⅴ类综合排序值和平均隶属度值均较低,抗低温能力最弱。
| 聚类Cluster | |||||
| Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅴ | |
| 综合排序值Comprehensive score | 3.87 | 3.32 | 2.87 | 2.32 | 1.77 |
| 平均隶属度值Average of SF | 0.57 | 0.49 | 0.37 | 0.31 | 0.20 |
各品种的聚类类别如图 9所示。良星619、丰德存麦1号、B07-4056、石H083-363、山农055843和良星99等6个品种聚在第Ⅰ类,其抗寒性最强;宿553、陕农509、A-9、中原6号、徐麦4036、舜麦1718和石麦19等7个品种聚在第Ⅱ类,其抗寒性强;尧麦16、C-44、山农05-066、冀麦585和石B05-7388等5个品种聚在第Ⅲ类,其抗寒性中等;偃展4110、B-33、B05-6507和石4185等4个品种聚在第Ⅳ类,其抗寒性弱;石06-6136和石优20这2个品种聚在第Ⅴ类,其抗寒性最弱。
| x1 | x2 | x3 | x4 | x5 | x6 | x7 | CS | ASF | |
| *表示相关性达显著性水平(P<0.05),**表示相关性达极显著水平(P<0.01); CS表示综合排序值,ASF表示平均隶属度值 | |||||||||
| x1 | 1.000 | ||||||||
| x2 | 0.230 | 1.000 | |||||||
| x3 | 0.422* | 0.479* | 1.000 | ||||||
| x4 | -0.027 | 0.327 | -0.068 | 1.000 | |||||
| x5 | 0.079 | -0.049 | -0.070 | -0.076 | 1.000 | ||||
| x6 | -0.169 | 0.204 | -0.043 | 0.025 | -0.223 | 1.000 | |||
| x7 | -0.091 | -0.015 | -0.093 | 0.339 | -0.244 | -0.224 | 1.000 | ||
| CS | -0.014 | 0.556* * | 0.393 | 0.691* * | -0.506* | 0.268 | 0.511* | 1.000 | |
| ASF | -0.149 | 0.640* * | 0.347 | 0.621* * | -0.503* | 0.404 | 0.412* | 0.970* * | 1.000 |
从相关关系来看(表 4),平均隶属度值和综合排序值与叶片相对可溶性糖含量、相对脯氨酸含量呈极显著正相关(P<0.01),与叶片相对POD活性呈显著正相关(P<0.05),与叶片相对MDA含量呈显著负相关(P<0.05)。二者与相对可溶性蛋白含量和相对SOD活性呈正相关,与相对电导率呈负相关。
从各品种的相对性状值来看(表 2),良星619(15号)叶片相对可溶性糖含量和相对脯氨酸含量均最高,其平均隶属函数值和综合排序值亦排在首位,抗寒性最高。石H083-363(3号)叶片相对MDA含量最低,相对SOD活性最高,其隶属函数值和综合排序值分别为0.604和3.763,抗寒性属于第I类;B07-4056(5号)叶片相对POD活性远高于其它品种,其隶属函数值和综合排序值分别为0.566和3.837,抗寒性亦属于第I类。石优20(2号)叶片相对MDA含量远高于其它品种,其隶属函数值和综合排序值分别为0.192和1.661,其抗寒性最弱;石06-6136(6号)叶片相对可溶性糖含量和相对SOD活性较低,而石4185(8号)叶片相对可溶性蛋白含量和相对脯氨酸含量均最低,这2个品种的抗寒性均弱,仅略高于石优20(2号)。
3 讨论拔节期是小麦快速生长时期,亦是对低温敏感的时期[14]。低温胁迫下,植物细胞发生一系列生理生化变化以适应和进行自我调节[4, 5, 6]。生物膜是植物细胞与外界环境之间发生物质交换的通道,各种逆境对植物细胞的影响首先作用于生物膜,常以叶片电导率作为鉴定细胞膜透性以及破坏程度的重要指标[7, 28]。本研究表明,拔节期低温胁迫后小麦叶片相对电导率较对照增加。因此,低温胁迫使小麦叶片生物膜透性增加并且膜受损程度加大。
低温胁迫下可溶性糖、可溶性蛋白和游离脯氨酸等渗透调节物质的含量发生变化,植物可通过调节渗透浓度来启动脱落酸的形成,诱发蛋白质的合成,增加抗寒性[9]。本研究表明,拔节期低温胁迫后,叶片可溶性糖含量和游离脯氨酸含量较对照增加,在一定程度上可以促进蛋白质的合成,但叶片中可溶性蛋白含量仍表现为低于对照。
低温胁迫下,植物体内产生大量的活性氧自由基,引发膜脂过氧化[8],产生膜脂过氧化产物MDA,且膜脂过氧化的程度随温度降低和胁迫时间延长而加重[9],同时植物体内还存在着SOD、POD等抗氧化酶系统以清除活性氧、降低逆境损伤[9, 10, 11]。本研究表明,拔节期低温胁迫下,小麦叶片SOD活性、POD活性和MDA含量均高于对照。SOD活性和POD活性的增加可以减轻低温对生物膜的迫害,说明植株对低温产生了一定的响应。但是MDA含量的增加显示低温胁迫后叶片膜脂过氧化程度仍然呈现增高趋势。
不同小麦品种抗低温能力差异很大[8, 14, 24, 26, 27, 30, 31]。 有研究表明,低温胁迫下小麦叶片MDA含量[8, 14]、可溶性蛋白含量[8]下降,电导率[29]、SOD活性[6, 29]、POD活性[6]、可溶性糖含量[14]的变化存在品种间差异或变化不明显。这主要是由于品种特性、胁迫温度强度、胁迫持续时间及小麦所处生育期不同而致。本研究表明,拔节期低温胁迫后,小麦叶片相对电导率、可溶性糖含量、游离脯氨酸含量、MDA含量、POD活性品种间极差值均高于对照,而可溶性蛋白含量和SOD活性品种间极差值低于对照,不同品种间各指标的增加(减少)量存在较大差异。计算各指标的相对性状值,则可以消除品种自身遗传特性的影响[8, 14, 15]。本研究表明,低温胁迫后,相对电导率的相对值以及相对脯氨酸含量较高,而相对可溶性蛋白含量较低。由相对性状值,通过隶属函数法和极点排序法,计算出各品种的平均隶属度值和综合排序值,并通过K-means聚类,将参试品种聚成了5类。其中,良星619抗低温能力最强,而石优20抗低温能力最弱。
有关小麦抗寒性的评价指标,已有研究并不完全一致。刘艳阳研究认为SOD活性、MDA含量可以作为小麦抗寒性的评价指标[6],高志强认为电导率和可溶性糖含量是衡量小麦抗寒性较好的指标[28]。本研究相关分析表明,叶片相对可溶性糖含量、相对脯氨酸含量、相对POD活性与相对MDA含量与平均隶属度值和综合排序值相关关系显著,认为这4个指标可以作为拔节期小麦抗寒性鉴定的主要评价指标。
本研究仅从叶片生理响应上分析了拔节期低温对小麦植株生长的影响,而小麦的抗寒性不仅包含自身的遗传因素和生理特征,更与外界环境因素密切有关。选育抗寒性强的品种、建立良种良法配套的高产栽培技术,提高小麦的抗寒综合能力,是目前小麦安全高效栽培生产中亟待解决的问题。
| [1] | Fang S B, Yang J J, Zhou G X. Change trend distributive characteristics of agrometeorological disasters in China in recent 30 years. Journal of Natural Disasters, 2011, 20(5): 69-73. |
| [2] | Wang J F, Zhu Y Y, Liu H P. Main agrometeorological disasters and their impact in recent 28 a in Henan. Meteorological and Environmental Sciences, 2007, 30(Suppl.): 9-11. |
| [3] | Cao H M, Shi Z M, Zhou X B, Lei P Z, Dong S G. A review on response of plant to low temperature and its cold resistance. Chinese Journal of Agrometeorology, 2010, 31(2): 310-314. |
| [4] | Gana J A, Sutton F, Kenefick D G. cDNA structure and expression patterns of a low-temperature-specific wheat gene tacr7. Plant Molecular Biology, 1997, 34: 643-650. |
| [5] | Minami A, Nagao M, Ikegami K, Koshiba T, Arakawa K, Fujikawa S, Takezawa D. Cold acclimation in bryophytes: low-temperature-induced freezing tolerance in Physcomitrella patens is associated with increases in expression levels of stress-related genes but not with increase in level of endogenous abscisic acid. Planta, 2005, 220: 414-423. |
| [6] | Liu Y Y, Li J Z, Chen L, Cui D Q. Effect of low temperature stress on peroxidation product of membrane lipids and activity of related enzymes in wheat seeding leaves. Journal of Triticeae Crops, 2006, 26(4): 70-73. |
| [7] | Wang J Y, Yu S Q. Studies on resistance and membrane lipid fluidity of various breeds of cold-resistant wheat. Biotechnology, 1998, 8(2): 28-30. |
| [8] | Wang S G, Wang Z L, Wang P, Wang H W, Li F, Huang W, Wu Y G, Yin Y P. Evaluation of wheat freezing resistance based on the responses of the physiological indices to low temperature stress. Acta Ecologica Sinica, 2011, 31(4): 1064-1072. |
| [9] | Li C Y, Chen S S, Xu W, Li D S, Gu X, Zhu X K, Guo W S, Feng C N. Effect of low temperature at seedling stage on Antioxidation enzymes and Cytoplasmic Osmoticum of leaves in wheat cultivar Yangmai 16. Acta Agronomica Sinica, 2011, 37(12): 2293-2298. |
| [10] | Allen R D. Dissection of oxidative stress tolerance using transgenic plants. Plant Physiology, 1995, 107: 1049-1054. |
| [11] | Wang X N, Fu L S, Li Z F, Sun Y L, Wang Y B, Liu C, Wang J W, Chen Y X. Morphogenesis and physiological basis in wheat cultivars with different levels of cold-resistance during cold acclimation and freezing period. Acta Agronomica Sinica, 2009, 35(7): 1313-1319. |
| [12] | Wang X X, Li S D, Dong H R, Gao Z H, Dai S S. The relationship between tomato varieties resistance and ABA and soluble sugar content. Acta Horticulturae Sinica, 1998, 25(1): 56-60. |
| [13] | Liu H Y, Zhu Z J, Lv G H, Qian Q Q. Study on relationship between physiological changes and chilling tolerance in grafted watermelon seedlings under low temperature stress. Scientia Agricultura Sinica, 2003, 36(11): 1325-1329. |
| [14] | Chen L, Wu S G, Li S M, Yin G H, Zhou L, Gu H M. Biochemical reaction and resistance analysis of jointing stage of winter wheat under low temperature stress. Acta Agriculturae Boreali-Sinica, 2001, 16(4): 42-46. |
| [15] | Yang M, Guo J Z. Evaluation of mulberry cold resistance based on the responses of the physiological indices to low temperature stress. Northern Horticulture, 2012,(4): 29-32. |
| [16] | Du Y J, Yu L, Sun J X, Lu W H. Comprehensive assessment of cold resistance of three Zoysia japonica varieties. Acta Prataculturae Sinica, 2008, 17(3): 6-16. |
| [17] | Xu N. Application of fuzzy comprehensive evaluation in the cold resistance identification of tea germplasms. Tea in Fujian, 1993, (3): 16-19. |
| [18] | Tang S Y, Yang L T, Li Y R. Response of different sugarcane genotypes to low temperature and evaluation for cold tolerance of sugarcane. Chinese Journal of Tropical Crops, 2012, 33(4): 635-641. |
| [19] | Zhang B Q, Yang L T, Li Y R. Comparison of physiological and biochemical characteristics related to cold resistance in sugarcane under field conditions. Acta Agronomica Sinica, 2011, 37(3): 496-505. |
| [20] | Zhang W E, Wang F, Pan X J. Comprehensive evaluation on cold hardiness of Vitis species by Subordinate Function (SF). Journal of Fruit Science, 2007, 24(6): 849-853. |
| [21] | Liu X M, Shang Q M, Zhang Z G. Low-temperature tolerance of pepper at germination stage and its evaluation method. Chinese Journal of Eco-Agriculture, 2005, 18(3): 521-527. |
| [22] | Liu X M, Shang Q M, Wang X L, Zhang Z G, Liu J C, Cui S M. Studies on low temperature tolerance evaluation method of different cucumber varieties at early stage. Journal of Inner Mongolia Agricultural University, 2010, 31(4): 154-160. |
| [23] | Ma W G, Cui H W, Li Y P, Zheng Y Y, Han R, Hu J. Effects of low temperature stress on physiological and biochemical characteristics of 20 tobacco cultivars and assessment of their cold tolerance at germination and seedling stages. Acta Agriculturae Zhejiangensis, 2011, 23(2): 232-238. |
| [24] | Li H S. The experiment principle and technique on plant Physiology and biochemistry. Beijing: Higher Education Press, 2000, 167-139, 184-185, 195-197, 261-263. |
| [25] | Shanghai Institute of Plant Physiology; Academia Sinica, Shanghai Institute of Plant Physiology. Modern Plant Physiology Laboratory Manual. Beijing: Science Press, 1999: 303. |
| [26] | Hou F L. Plant Physiology Experimental Course. Beijing: Science Press, 2004, 91. |
| [27] | Ren X L, Li J R. Changes of active oxygen and some physiological indexes in the processes of maturation and senescence of apricot fruits. Plant Physiogy Communications, 1991, 27(1): 34-36. |
| [28] | Gao Z Q, Zhang G H, Zhang A Z, Miao G Y. The low temperature reaction of the different wheat varieties. Journal of Shanxi Agricultural University, 2002, 22(2): 109-112. |
| [29] | Yi Y, Zhang Y L, Guo Z F, Bai L P, Qiao X, Hou L B. Physiological responses of winter wheat's leaves to low temperature stress. Acta Agriculturae Boreali-Sinica, 2013, 28(1): 144-148. |
| [30] | Yu J, Zhang L, Cang J, Hao Z B, Wang D J. Research progress of cold resistance in winter wheat. Journal of Northeast Agricultural University, 2008, 39(11): 123-127. |
| [31] | Feng S W, Dong N, Hu T Z, Li G, Li X H, Ru Z G. Analysis on the integrated winter hardness of different wheat varieties. Journal of Henan Institute of Science and Technology, 2010, 38(3): 4-7. |
| [1] | 房世波, 阳晶晶, 周广胜. 30年来我国农业气象灾害变化趋势和分布特征. 自然灾害学报. 2011, 20(5): 69-73. |
| [2] | 王记芳, 朱业玉, 刘和平. 近28a河南主要农业气象灾害及其影响. 气象与环境科学, 2007, 30(增刊):9-11. |
| [3] | 曹慧明,史作民,周晓波,雷鹏志, 董生刚. 植物对低温环境的响应及其抗寒性研究综述. 中国农业气象, 2010, 31(2):310-314. |
| [6] | 刘艳阳,李俊周,陈磊,崔党群. 低温胁迫对小麦叶片细胞膜脂过氧化产物及相关酶活性的影响. 麦类作物学报, 2006, 26(4): 70-73. |
| [7] | 王纪元, 喻尚其. 不同抗寒性小麦品种的抗性与膜脂流动性的研究. 生物技术, 1998, 8(2): 28-30. |
| [8] | 王树刚, 王振林, 王 平,王海伟,李府,黄玮,武玉国,尹燕枰. 不同小麦品种对低温胁迫的反应及抗冻性评价. 生态学报, 2011, 31(4): 1064-1072. |
| [9] | 李春燕,陈思思,徐雯, 李东升, 顾骁, 朱新开, 郭文善, 封超年. 苗期低温胁迫对扬麦16叶片抗氧化酶和渗透调节物质的影响. 作物学报, 2011, 37(12): 2293-2298. |
| [11] | 王晓楠, 付连双, 李卓夫, 孙艳丽,王玉波,刘灿,王金伟,陈禹兴. 低温驯化及封冻后不同抗寒性小麦品种的形态建成及生理基础分析. 作物学报, 2009, 35(7): 1313-1319. |
| [12] | 王孝宣, 李树德, 东惠茹, 高振华, 戴善书. 番茄品种耐寒性与ABA和可溶性糖含量的关系. 园艺学报, 1998, 25(1): 56-60. |
| [13] | 刘慧英, 朱祝军, 吕国华, 钱琼秋. 低温胁迫下西瓜嫁接苗的生理变化与耐冷性关系的研究. 中国农业科学, 2003, 36(11): 1325-1329. |
| [14] | 陈龙,吴诗光, 李淑梅,殷贵鸿, 周琳, 古红梅. 低温胁迫下冬小麦拔节期生化反应及抗性分析. 华北农学报. 2001, 16(4): 42-46. |
| [15] | 杨梅,郭军战. 不同果桑品种对低温胁迫的反应及抗寒性评价. 北方园艺, 2012, (4): 29-32. |
| [16] | 杜永吉,于磊,孙吉雄,鲁为华. 结缕草3个品种抗寒性的综合评价. 草业学报,2008, 17(3): 6-16. |
| [17] | 许宁. 模糊综合评判在茶树种质资源抗寒性鉴定中的应用研究.福建茶叶, 1993, (3): 16-19. |
| [18] | 唐仕云,杨丽涛,李杨瑞. 不同甘蔗基因型对低温胁迫的响应及其耐寒性评价. 热带作物学报. 2012, 33(4): 635-641. |
| [19] | 张保清, 杨丽涛, 李杨瑞. 自然条件下甘蔗品种抗寒生理生化特性的比较. 作物学报, 2011, 37(3): 496-505. |
| [20] | 张文娥, 王飞, 潘学军. 应用隶属函数法综合评价葡萄种间抗寒性. 果树学, 2007, 24(6): 849-853. |
| [21] | 刘雪梅, 尚庆茂, 张志刚. 辣椒不同品种种子萌芽期耐低温性及评价方法研究. 中国生态农业学报, 2005, 18(3): 521-527. |
| [22] | 刘雪梅, 尚庆茂, 王晓黎, 张志刚, 刘杰才, 崔世茂. 黄瓜不同品种耐低温性早期评价方法研究. 内蒙古农业大学学报, 2010, 31(4): 154-160. |
| [23] | 马文广,崔华威,李永平,郑昀晔, 韩瑞, 胡晋. 20个烟草品种低温逆境下发芽和苗期生理生化特性的变化及耐寒性评价. 浙江农业学报. 2011, 23(2): 232-238. |
| [24] | 李合生. 植物生理生化实验原理和技术. 北京: 高等教育出版社, 2000, 167-139, 184-185, 195-197, 261-263. |
| [25] | 中国科学院上海植物生理研究所, 上海植物生理学会. 现代植物生理学实验指南. 北京: 科学出版社, 1999: 303. |
| [26] | 侯福林. 植物生理学实验教程. 北京: 科学出版社, 2004, 91. |
| [27] | 任小林,李嘉瑞. 杏果实成熟衰老过程中活性氧和几种生理指标的变化(简报). 植物生理学通讯, 1991, 27(1): 34-36. |
| [28] | 高志强, 张国红, 张爱芝, 苗果园. 不同小麦品种对低温的生理反应研究. 山西农业大学学报, 2002, 22(2): 109-112. |
| [29] | 衣莹, 张玉龙, 郭志富, 白丽萍, 乔雪, 侯立白. 冬小麦叶片对低温胁迫的生理响应. 华北农学报, 2013, 28(1): 144-148. |
| [30] | 于晶, 张林, 苍晶, 郝再彬, 王多佳. 冬小麦抗寒性的研究进展. 东北农业大学学报, 2008, 39(11): 123-127. |
| [31] | 冯素伟, 董娜, 胡铁柱, 李淦, 李笑慧, 茹振钢. 不同小麦品种抗低温能力比较. 河南科技学院学报, 2010, 38(3): 4-7. |