 2014, Vol. 34
2014, Vol. 34文章信息
- 孙忠林, 王传宽
- SUN Zhonglin, WANG Chuankuan
- 森林生态系统可溶性碳和颗粒碳通量
- Dissolved and particulate carbon fluxes in forest ecosystems
- 生态学报, 2014, 34(15): 4133-4141
- Acta Ecologica Sinica, 2014, 34(15): 4133-4141
- http://dx.doi.org/10.5846/stxb201302270309
-
文章历史
- 收稿日期:2013-2-27
- 修订日期:2014-4-11






2. 通化师范学院, 通化 134002
2. Tonghua Normal University, Tonghua 134002, China
The DC and PC in a forest ecosystem mainly consist of dissolved organic C (DOC), dissolved inorganic C (DIC), particulate organic C (POC), black C and carbonate C, which fundamentally originate from net primary production of the ecosystem. DOC, an important active pool of C and nutrients, is mainly composed of carbohydrates, long-chain aliphatic compounds, aromatic compounds, proteins, and low molecular weight organic acids. The DOC content is correlated positively with concentrations of proteins and carbohydrates, but negatively with concentrations of carboxyl C. The DOC in soil solution is mainly derived from plant litter, soil humus, microbial debris, root exudates, and ectomycorrhizal mycelium. DIC consists of dissolved inorganic carbonate, dissolved bicarbonate, dissolved carbonate and dissolved carbon dioxide. It activates soil nutrient elements and thus enhances plant growth. POC is the major form of PC in forest soils that includes polymeric gel, organic fiber, cell fragments, microbial biomass (such as algae and bacteria), and polymerized DOC. Both DC and PC are labile C pools in the forest soil system, which enter the ecosystem from atmospheric deposition, throughfall and litterfall, and leave the ecosystem via soil respiration, lateral transport and seepage. At a small local scale, fluxes of DC and PC are mainly influenced by such biotic factors as root exudates, fine root decomposition and microbial turnover. At a large regional scale, they are jointly affected by soil and vegetation characteristics, coupling effects of ecological processes, climatic conditions and global changes. Future research priorities in this field should be: (1) to explore the factors controlling fluxes of DC and PC in forest ecosystems on various spatio-temporal scales, their potential coupling relationships and underlying mechanisms; (2) to understand interactions and transformation between DC/PC and other C fluxes in forest ecosystems, and explore potential stoichiometric relationships between DC/PC and other nutrients in the systems and (3) to explore effects of global changes on fluxes of DC and PC in forest ecosystems, particularly focusing on the effects of extreme disturbances, such as forest management, wildfire, drought, flooding, ice and frozen, and freeze-thaw cycles.
森林生态系统是陆地生态系统中面积最大、最复杂的自然生态系统[1]。在全球变化的背景下,森林的生态服务功能,特别是其碳汇功能日益凸显[2]。森林碳汇功能常采用生态系统净生产力(NEP)或生态系统净碳交换(NEE)等指标来量化。近20年来,全球和区域通量网络发展迅速,已经基本实现了连续监测全球主要陆地生态系统与大气圈之间的NEE[3],为定量评价陆地生态系统碳收支、构建和校验碳循环模型提供了重要的实测数据[3, 4]。然而,陆地生态系统在与大气圈进行碳交换的同时,也与水域生态系统(溪流、湖泊、海洋等)进行着物质和能量的交换。这部分碳通量通过液态形式进出森林生态系统,是目前常规碳计量方法(如测树学方法、涡动协方差法等)无法测定的,因而在森林固碳功能的评价和模型预测中常被忽略。针对碳循环过程发生在多个系统之间、涉及的时空尺度较大等特点,Chapin等[5]建议,采用如下生态系统净碳平衡(NECB)的概念可更全面地量化评价陆地生态系统碳收支:
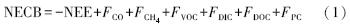
式中,FCO、FCH4、FVOC、FDIC、FDOC、FPC分别表示一氧化碳、甲烷、挥发性有机碳、可溶性无机碳(DIC)、可溶性有机碳(DOC)和颗粒碳(PC)通量。
DC和PC是森林生态系统碳收支的重要组分,但由于认识不足、测定困难等原因,在以往的森林碳汇研究中常被忽略[5],因而增加了森林固碳功能评价和模型预测的不确定性。例如,有研究认为,全球碳循环研究中的“碳失汇”现象很可能与北半球森林生态系统DC和PC流失有关[6]。另外,森林生态系统DC和PC通量的量化可用于校正测树学方法、涡动协方差法等常规方法测得的森林碳汇结果[5]。为此,本文对森林生态系统DC和PC通量的组成、作用、相关生态过程及其影响因子等进行了综述,并提出该领域应该优先考虑的研究问题,以便全面系统地理解森林碳循环过程,减少森林碳收支计量评价和预测中的不确定性。
1 DC和PC的组成与作用DC主要由DOC和DIC组成。DC和PC是森林土壤的重要活性碳库[7],其中一部分被微生物分解,以气态形式回归到大气或成为微生物的组分[8],另一部分通过土壤水分作侧向和纵向移动,伴随着地表径流、壤中流和地下水输出森林生态系统。据Meybeck[9]估测,全球通过河流从陆地输入海洋的总碳量为542 Tg C/a,其中DOC、DIC和PC分别占37%、45%和18%。
1.1 DOCDOC通常指能够通过0.45 μm滤器的大小和结构不同的水溶性有机分子[10],一般由碳水化合物、长链脂肪族化合物、芳香族化合物、蛋白质、低分子量有机酸等组成,其含量与蛋白质、碳水化合物呈正相关,与羧基碳呈负相关[11, 12]。土壤溶液中的DOC主要来源于植物凋落物、土壤腐殖质、微生物残体、根系分泌物[13]、外生菌根等[14],其中仅能化学鉴别出一小部分低分子DOC(如有机酸、糖类、氨基酸等),而多数成分为组成复杂、分子量高、无法命名的腐殖类物质。DOC通常占总初级生产量(GPP)的比例较小,但是与NEP相比则不可忽视。例如,Gielen 等[15]报道,温带欧洲赤松林DOC通量仅占GPP的0.8%,却占NEP的17%。
DOC是森林生态系统中的活性碳库和重要养分库;它随着河川径流输入海洋,成为海洋生态系统中DOC的重要组分[16]。DOC除了作为NECB组分之外,在陆地和水域生态系统中均具有促进矿物风化、为微生物生长和生物分解提供能量、提高生态系统养分有效性[17, 18, 19]、降低污染物毒性[20, 21]等作用。
1.2 DICDIC是指全部或部分溶解于水的无机碳酸盐、碳酸氢盐、碳酸及二氧化碳的总称。土壤中产生的CO2可部分溶解于水,导致土壤水分中CO2过饱和,再通过河川径流转运后重新释放到大气中[18]。土壤微生物在分解利用DOC的同时,可以将其转变为DIC,并通过径流输出该生态系统,这部分碳通量可高达土壤溶液DOC总量的40%[19]。另外,DIC也是一些水分充足的森林生态系统(如红树林)与相邻水域进行碳交换的主要形式[22]。
土壤溶液中CO2过多,可使土壤酸化,导致土壤中的阳离子(特别是铝离子)活化。例如,Amirbahman等[23]报道,DIC浓度升高可使土壤溶液中的金属及营养元素活化,从而引起森林生态系统地表水水质恶化。另外,植物根系可吸收土壤中的DIC,通过茎干液流运至叶片,成为光合作用的底物[24]。虽然这部分DIC比叶片从大气中吸收CO2作为光合作用底物的量小得多,但对植物幼嫩茎干和细根的生长具有重要意义[24]。
1.3 PCPC的定义在文献中有分歧。较早研究常以0.7 μm作为划分PC与DC的临界值[25];近期研究多以0.45 μm为标准[26];也有人建议视具体情况而定[27]。从全球碳预算的可比性以及概念统一的角度考虑,Michalzik和Stadler[28]建议将PC的上限定为2 mm;而Le Mellec等[29]则提出将PC的上限定为500 μm。通常,人们把PC分成三大类:黑碳(BC)、颗粒有机碳(POC)和碳酸盐碳(CC)[26]。
BC是化石燃料和生物量不完全燃烧而生成的高芳香化结构的含碳颗粒物,其碳含量高达60%以上[30]。在北方森林中,林火作为一个重要的生态干扰因子,在将碳以气体形式释放到大气而成为重要碳源的同时,又将有机碳转变成难降解的BC而成为常被人们忽略的碳汇。1977—1990年期间,北半球由于林火造成的碳源约为0.13 Pg C/a[31],碳汇则波动在0.05—0.27 Pg C/a之间[32]。
POC由聚合胶体、有机纤维、细胞片段及微生物生物质(如藻类与细菌)组成,也可由DOC聚合而成[33]。POC是土壤PC的主要形式,无论在土壤孔隙还是在溪流生态系统中,其通量都仅次于DOC[26]。其实,DOC和POC很难确切地划分,主要是因为两者在特定条件下可相互转化,因此简单地将DOC定义为胶体状态、POC为悬浮状态的看法可能不妥。
CC来源于土壤碳酸盐水解作用和裸露在大气中的土壤碳酸盐裂解、淋溶和风化作用,主要存在于土壤大颗粒有机质之间。CC通量与BC和POC通量相比很小,因此在许多研究中被忽略[34],但随土壤溶液酸化的加强,其溶解的比例会增大。
2 DC和PC通量的相关生态过程以森林生态系统的土壤子系统为中心,DC和PC通量主要包括输入、转化和输出3个基本生态过程(图 1),它们既有相对独立性又相互影响。

|
| 图 1 森林生态系统可溶性碳和颗粒碳通量相关的生态过程 Fig. 1 The ecological processes associated with dissolved carbon and particulate carbon fluxes in forest ecosystems |
在森林生态系统中,虽然大气碳沉降的绝对数量不大,但随着化石燃料利用的增加和人类活动的加剧而有增加趋势。Jones等[8]估测北威尔士大气碳湿沉降约为1 g C m-2 a-1,仅相当于当地土壤呼吸年通量的0.67%。在冬季取暖以煤炭为主的温带和北方地区,含碳物质的不完全燃烧会在大气中形成黑碳气溶胶,并以干、湿沉降的形式输入森林生态系统。尤其是在林火频繁发生的北方森林中,生物量和土壤有机碳在火烧过程中部分转换为BC粉尘颗粒,可占植物燃烧产生的BC总量的20%左右[35]。在一些地表扰动较大、植被覆盖率较低的地区,土壤微粒可进入大气形成胶体,在一定条件下以干、湿沉降的形式返回生态系统。
穿透雨是指大气降雨穿过林冠层到达土壤的那一部分降雨。大气降雨一方面将部分大气粉尘颗粒带回陆地,另一方面将植被表面的DC和PC“冲刷”带回土壤,这种“淋洗”作用在森林生态系统中尤为突出。例如,Le Mellec等[29]报道,德国北部云杉林生态系统中穿透雨中总有机碳平均浓度高达18 mg/L,其中DOC和POC分别占84%和16%,DOC和POC通量分别为8.7 g C m-2 a-1和1.7 g C m-2 a-1。
森林凋落物和腐殖质是土壤DOC最重要的“源”,特别是新近凋落物可将大量DOC释放到土壤中[36]。然而,凋落物和腐殖质对土壤DOC的相对贡献率难以量化。凋落物和腐殖质的分解过程和淋溶作用影响着DC和PC向土壤的输入和转移,从而改变植物根系动态和土壤生物地球化学循环过程。虽然对凋落物DOC输入过程的主导驱动因素(生物因素或物理化学因素)还有争议,但凋落物量的增减显著地影响土壤DOC的输入量[37]。此外,由于大气碳沉降和穿透雨DC和POC输入的影响,森林地被物滤出液中DOC量通常与降雨和穿透雨中的DOC量呈显著正相关[38]。
2.2 转化土壤中DC和PC的转化过程很复杂,相关机理研究不多,多数研究将土壤视为“黑箱。”事实上,任何影响土壤有机碳含量的生态过程均影响土壤DC和PC的转化,其中主要包括根系分泌、细根分解、微生物周转等。
根系分泌过程是根际沉积的一部分,是植物直接向土壤释放DOC的重要过程。根系分泌物主要包括可溶性糖、低分子有机酸、氨基酸、酚类、大分子粘液、蛋白质等。许多根系分泌物具有他感作用,影响植物对环境的适应性及群落演替。另外,植物根系能借助于外生菌根菌丝体增强向土壤分泌DOC的能力[14]。
虽然细根生物量占森林生态系统总生物量的比例很小,但是细根生产量对净初级生产力的贡献率很高(33%—67%)[39]。细根生产量大、寿命短、分解快,在分解过程中不断向土壤释放非结构性碳水化合物和疏水酸,大大地增加了土壤有机碳的输入量[40]。
土壤微生物具有分解有机质和活性养分源的双重作用:既催化土壤有机质的分解而导致DOC的大量释放,又通过其自身死亡及残体裂解而成为土壤有机碳的重要活性组分。土壤微生物生物量碳仅占土壤总有机碳量的1%—3%,但其高周转率对土壤有机质的转化过程影响显著,尤其是植物外生菌根菌丝体是土壤微生物生物量的重要组分[14]。
2.3 输出土壤DC和PC主要通过土壤呼吸、侧向运输和渗透流失的方式输出森林生态系统(图 1)。土壤呼吸是森林生态系统中第二大碳通量(仅次于GPP)[41],其中异养呼吸是土壤有机质的主要输出方式。Wang等[42]采用挖壕法将我国主要温带森林土壤呼吸分离成根际呼吸和异养呼吸两部分,发现土壤异养呼吸占土壤呼吸年通量的17%—48%,且与土壤表层有机碳含量呈显著正相关。此外,土壤微生物生物量常与净初级生产力关系密切,增加凋落物、细根等新鲜有机质也会加快土壤异养呼吸速率[41]。
在地形起伏的森林生态系统中,水分在重力和坡度的双重作用下形成侧向径流(地表径流和壤中流)和渗流。由于森林土壤具有孔隙度大、渗透性高的特点,因此很少能观察到形成大量地表径流,因而以地表径流的方式输出的DC和PC较少。
DC和PC主要通过纵向的渗透作用最终以径流的方式输出森林生态系统,在母质层致密、母岩层透水性差的情况下尤为突出。例如,美国Hubbard Brook森林生态系统因有不透水岩层存在,其DC和PC以溪流输出为主[43]。由于土壤孔隙及其吸附作用,渗流以DC为主。降雨量越大、土壤孔隙度越大,径流DC输出量也越大。据Schmidt等[44]估测,在年降水量较高的亚热带地区,日本扁柏林的DOC通量波动在819—962 kg C hm-2 a-1之间;主要通过渗透输出的森林土壤滤出液DOC通量和渗流DOC通量分别为962 kg C hm-2 a-1和478 kg C hm-2 a-1。
3 DC和PC通量的影响因子DC和PC通量的受控因素很多[13],主要包括:植被与土壤特性[38, 45]、生态过程耦联关系[13, 46]、气候条件[47]、全球变化[48, 49]等。
3.1 植被与土壤特性植被是土壤有机质的最重要来源,其中细根分泌物、凋落物输入及其分解速率直接影响森林土壤的碳储量及其周转[37, 50]。Badri等[50]报道,植物幼苗将光合作用固定的有机物的30%—40%用于根系分泌,但根系分泌物的质和量与植物的种类、年龄等因素有关。Wang等[51]报道,我国亚热带森林中阔叶林的有机碳和DC含量均高于针叶林。Leff等[37]发现,热带森林凋落物的短期(两年)增减显著地影响土壤DOC、微生物生物量和细根生物量。此外,花粉沉降、昆虫排泄物、大气干湿沉降是森林冠层DC和PC的重要来源,因此,树种组成、季相变化等植被结构特征也会影响DC和PC浓度及其动态[38]。
土壤质地、孔隙度、温度、含水量、pH值、微生物丰度及活性等因素可影响DC和PC通量及其在森林生态系统中的利用率[17, 45, 52, 53]。Christ等[52]发现,红皮云杉林地被物DOC浓度随温度升高而呈指数型增长,其温度敏感性(Q10)随土壤含水量增加也有增加趋势。Bader和Krner等[54]的控制实验表明,土壤含水量增加可使土壤DC的渗透量增大,从而增加DC和PC的输出通量。Andreasson等[17] 的研究结果显示,两种不同的山毛榉林地上植被对DOC的影响不大,但其土壤微生物对土壤滤出液和底物溶液的利用效率的影响差异显著。此外,DOC通量还受土壤pH值影响,反之又能影响土壤pH值[53]。
3.2 生态过程耦联关系森林生态系统的结构和功能、功能过程之间相关联,形成错综复杂的耦联关系,从而影响DC和PC通量,其中水循环与碳循环耦联尤为紧密。水循环影响森林生态系统的光合作用、呼吸作用以及碳沉积[55],从而显著地影响森林碳循环过程。例如,Buckingham等[56]对11个森林样地的DOC组成及其周转的时空格局研究表明,土壤DOC通量与水通量相关显著。
土壤可溶性有机质(DOM)是微生物的底物和能量来源,包括DOC、可溶性有机氮(DON)、可溶性有机磷(DOP)[13]等,三者在生物地球化学循环中相互关联,因此在物质转化、转移及运输过程中往往相耦联[57]。这种有机物之间的耦合关系通常在C/N上有所体现[58],因此DOC流失常伴随着DON流失[38],并与C/N呈正相关[59, 60]。Lutz等[46]的间接碳控制假说指出,微生物好分解富氮的DOM,但当氮不受限制时,微生物的数量主要受DOC量的影响。DOC作为溪流等异养型生态系统的重要碳源,其浓度会影响微生物对其它物质的代谢速率,从而影响N、P动态[61];而N、P又是微生物细胞的重要组分,其丰度又会影响微生物的数量和组成,进而影响微生物种群之间的竞争关系,最终影响DOC的分解和吸收[62]。由于影响DOC的释放和存留的因子未必与DON、DOP直接相关,因此杨玉盛等[63]建议将三者的动态区别对待,分别研究三者与植物的营养关系。
3.3 气候条件DC和PC的运输动力主要来自于水分势能,因此,降水的时空格局直接影响DC和PC通量的相关生态过程。一般来说,DC和PC的溪流输出通量与降水特征密切相关,特别是在土壤含水量接近饱和时,强降雨或春季融雪径流期间DC和PC的输出通量急剧增加。Clark等[64]对泥炭土的室内控制实验结果表明,虽然高温少雨对DOC的产生有积极作用,但这种作用也可能被其它过程所掩盖。例如,干旱条件下由SO2-4引起的土壤酸化可以限制DOC的生产[65]。Couture等[47]报道,1989—2006年期间加拿大温带和北方地区河流的DOC浓度以0.05 mg L-1 a-1的速率升高;进一步的模型分析发现,DOC浓度升高与气候参数(如降水、空气温度、辐射强度)密切相关。
低温[66, 67]以及冻融交替事件[68]会影响森林土壤碳循环及其相关过程,从而影响DC和PC的输出。Buckingham等[56, 67]报道,冬季DOC输出量占其年输出总量的75%。冻融交替事件影响微生物及其种群组成、根系周转、土壤结构、有机质分解等,进而影响冻融后DC和PC通量,但作用强度和机制尚无定论。例如,gren等[69]发现,高纬度和高海拔地区冬季温度越低,持续时间越漫长,则翌年春夏季的DC和PC通量越大;而Monson等[70]则得出相反结论;也有研究[67]报道冻融对土壤DOC渗透损失影响不大的结论。导致这种相悖结论的原因还不清楚。冻融交替事件特性(冻融强度、频度、持续时间等)不同,对土壤碳循环过程的影响机制不一,因而对DC的影响也会有差异[66]。Schmitt等[68]在室内控制实验中发现,频繁冻融事件对土壤中木质素的影响不显著,但磷脂脂肪酸含量降低;糖类含量减少,真菌菌丝破坏大,而微生物生物量总体上变化不显著。Feng等[71]研究表明,冻融过程中底物变化会使细菌与真菌之间的竞争关系发生改变,从而改变微生物群落组成,进而影响土壤的碳代谢过程。
3.4 全球变化由于人为活动加剧,致使全球CO2等温室气体浓度升高、平均气温升高、降水时空格局改变、极端干旱和洪涝事件频发、氮沉降加剧等全球变化,使森林生态系统的结构和功能过程发生深刻的变化,从而通过上述因子影响DC和PC通量。全球CO2浓度升高导致生态系统的DOC浓度升高,但两者之间的关系是非线性的[48]。例如,在过去20年期间,世界范围内地表水的DOC浓度明显增加,这与气候变化和大气中硫酸盐沉降的降低有关[49]。大气氮沉降促使森林地上和地下生物量增加,潜在地增加了DOC和POC的生产底物,使其在土壤中的浓度增加。多代营造人工纯林不但使土壤DOM下降,而且使其它养分也随之下降,造成地力衰退[72]。可见,在全球变化背景下,DC和PC通量的继续增大会严重影响森林生态系统碳收支,甚至改变森林的碳源汇功能[73]。
4 结语与展望DC和PC通量作为森林生态系统碳收支的重要组分,在森林固碳功能的评价和模型预测中具有重要意义,但由于认识不足、测定困难等原因而在森林碳汇研究中常被忽略。近年来,国外的相关研究从DC与PC运输过程的描述[7, 11, 38]逐渐转向阐明其周转过程及机理[13, 14, 74];然而,国内对DC和PC通量的系统研究较少,研究侧重于森林经营管理对DOM的影响[72]、退化生态系统修复过程中DOM的变化及其与土壤养分和微生物的关系[75] 等。至今为止,对DC和PC通量的驱动因子及其交互作用、影响程度及作用机理还不清楚,阻碍了DC和PC通量模型的构建和预测。例如,非生物因子(如温度、水文)的变化在一定程度上掩盖了生物因素(如植被)对DC和PC的影响[76];土壤DOC的吸附与去吸附作用在一定程度上会影响微生物的矿化作用,进而影响DOC周转,但影响程度不详。野外试验与室内控制实验、不同时空尺度(如样地、集水区、流域尺度)对DC和PC通量的测定结果不符,甚至得出相悖的结论[13],这很可能与不同试验条件下众多的影响因子和复杂的耦联关系有关,从而阻碍了不同研究结果的比较和整合。
为了全面系统地理解森林碳循环过程、减少森林碳收支计量评价和预测中的不确定性,有关森林生态系统DC和PC通量的研究应该优先考虑如下几个方面:(1)针对不同的时间和空间尺度,探索森林生态系统DC和PC通量的控制因子及其耦联关系,揭示其中的驱动机理,为构建DC和PC通量以及森林碳循环机理模型提供可靠的数据和理论基础。由于DC和PC通量影响因素的多样性以及森林生态系统在时空尺度上的高异质性,预测DC和PC通量的时空动态将是一个严峻的挑战[77]。(2)探索DC和PC与其它森林生态系统碳组分的相互关系和转化机理,阐明DC和PC通量与其它养分之间潜在的生态化学计量关系[46, 58, 59, 60, 61, 78]。例如,DOC可被微生物利用,也可随土壤理化条件的变化而沉积或矿化成无机碳[17, 45];DOC可被微生物呼吸输出,也可在其它系统中沉积或重新合成难于分解的芳香族化合物。这对于评价预测森林碳汇功能很重要。(3)随着人为活动和全球变化的加剧,森林生态系统的结构和功能过程将发生深刻变化,不仅会影响森林生态系统DC和PC通量,甚至会改变森林的分布格局及其碳源汇功能。因此,探索全球变化,特别是人类活动(如森林经营)和极端干扰事件(诸如林火、旱涝、冰冻、冻融交替等)对森林生态系统碳循环的影响,无疑是该领域的一个重要研究命题。
| [1] | Woodwell G M, Whitaker R H, Reiners W A, Likens G E, Delwiche C C, Botkin D B. The biota and the world carbon budget. Science, 1978, 199(4325): 141-146. |
| [2] | Pan Y, Birdsey R A, Fang J, Houghton R, Kauppi P E, Kurz W A, Phillips O L, Shvidenko A, Lewis S L, Canadell J G, Ciais P, Jackson R B, Pacala S W, McGuire A D, Piao S, Rautiainen A, Sitch S, Hayes D. A large and persistent carbon sink in the world's forests. Science, 2011, 333(6045): 988-993. |
| [3] | Friend A D, Arneth A, Kiang N Y, Lomas M, Ogée J, Rödenbeck C, Running S W, Santaren J D, Sitch S, Viovy N, Woodward F I, Zaehle S. FLUXNET and modelling the global carbon cycle. Global Change Biology, 2006, 13(3): 610-633. |
| [4] | Owen K E, Tenhunen J, Reichstein M, Wang Q, Falge E, Geyer R, Xiao X M, Stoy P, Ammann C, Arain A, Aubinet M, Aurela M, Bernhofer C, Chojnicki B H, Granier A, Gruenwald T, Hadley J, Heinesch B, Hollinger D, Knohl A, Kutsch W, Lohila A, Meyers T, Moors E, Moureaux C, Pilegaard K, Saigusa N, Verma S, Vesala T, Vogel C. Linking flux network measurements to continental scale simulations: ecosystem carbon dioxide exchange capacity under non-water-stressed conditions. Global Change Biology, 2007, 13(4): 734-760. |
| [5] | Chapin F S, Woodwell G, Randerson J T, Rastetter E B, Lovett G M, Baldocchi D D, Clark D A, Harmon M E, Schimel D S, Valentini R, Wirth C, Aber J D, Cole J J, Goulden M L, Harden J W, Heimann M, Howarth R W, Matson P A, McGuire A D, Melillo J M, Mooney H A, Neff J C, Houghton R A, Pace M L, Ryan M G, Running S W, Sala O E, Schlesinger W H, Schulze E D. Reconciling carbon-cycle concepts, terminology, and methods. Ecosystems, 2006, 9(7): 1041-1050. |
| [6] | Willey J D, Kieber R J, Eyman M S, Avery G B. Rainwater dissolved organic carbon: concentrations and global flux. Global Biogeochemical Cycles, 2000, 14(1): 139-148. |
| [7] | Aravena R, Wassenaar L I, Spiker E C. Chemical and carbon isotopic composition of dissolved organic carbon in a regional confined methanogenic aquifer. Isotopes in Environmental and Health Studies, 2004, 40(2): 103-114. |
| [8] | Jones D L, Hughes L T, Murphy D V, Healey J R. Dissolved organic carbon and nitrogen dynamics in temperate coniferous forest plantations. European Journal of Soil Science, 2008, 59(6): 1038-1048. |
| [9] | Meybeck M. Riverine transport of atmospheric carbon: sources, global typology and budget. Water, Air, & Soil Pollution, 1993, 70(1/4): 443-463. |
| [10] | Herbert B, Bertsch P. Characterization of dissolved and colloidal organic matter in soil solution: a review. Carbon Forms and Functions in Forest Soils, 1995: 63-88. |
| [11] | Candler R, Zech W, Alt H G. Characterization of water-soluble organic substances from a typic Dystrochrept under spruce using GPC, IR, 1H NMR, and 13C NMR spectroscopy. Soil Science, 1988, 146(6): 445-452. |
| [12] | Liang B C, Mackenzie A E, Schnitzer M, Monreal C M, Voroney P R, Beyaert R P. Management-induced change in labile soil organic matter under continuous corn in eastern Canadian soils. Biology and Fertility of Soils, 1997, 26(2): 88-94. |
| [13] | Kalbitz K, Solinger S, Park J H, Michalzik B, Matzner E. Controls on the dynamics of dissolved organic matter in soils: a review. Soil Science, 2000, 165(4): 277-304. |
| [14] | Högberg M N, Högberg P. Extramatrical ectomycorrhizal mycelium contributes one-third of microbial biomass and produces, together with associated roots, half the dissolved organic carbon in a forest soil. New Phytologist, 2002, 154(3): 791-795. |
| [15] | Gielen B, Neirynck J, Luyssaert S, Janssens I A. The importance of dissolved organic carbon fluxes for the carbon balance of a temperate Scots pine forest. Agricultural and Forest Meteorology, 2011, 151(3): 270-278. |
| [16] | Hansell D A, Kadko D, Bates N R. Degradation of terrigenous dissolved organic carbon in the western Arctic Ocean. Science, 2004, 304(5672): 858-861. |
| [17] | Andreasson F, Bergkvist B, Baath E. Bioavailability of DOC in leachates, soil matrix solutions and soil water extracts from beech forest floors. Soil Biology & Biochemistry, 2009, 41(8): 1652-1658. |
| [18] | Davidson E A, Figueiredo R O, Markewitz D, Aufdenkampe A K. Dissolved CO2 in small catchment streams of eastern Amazonia: a minor pathway of terrestrial carbon loss. Journal of Geophysical Research, 2010, 115(G4): G04005. |
| [19] | Shibata H, Mitsuhashi H, Miyake Y, Nakano S. Dissolved and particulate carbon dynamics in a cool-temperate forested basin in northern Japan. Hydrological Processes, 2001, 15(10): 1817-1828. |
| [20] | Antoniadis V, Tsadilas C D, Stamatiadis S. Effect of dissolved organic carbon on zinc solubility in incubated biosolids-amended soils. Journal of Environment Quality, 2007, 36(2): 379-385. |
| [21] | Berndt M E, Bavin T K. Methylmercury and dissolved organic carbon relationships in a wetland-rich watershed impacted by elevated sulfate from mining. Environmental Pollution, 2011, 161: 321-327. |
| [22] | Bouillon S, Borges A V, Castaneda-Moya E, Diele K, Dittmar T, Duke N C, Kristensen E, Lee S Y, Marchand C, Middelburg J J, Rivera-Monroy V H, Smith T J Ⅲ, Twilley R R. Mangrove production and carbon sinks: a revision of global budget estimates. Global Biogeochemical Cycles, 2008, 22(2): GB2013, doi:10.1029/2007GB003052. |
| [23] | Amirbahman A, Holmes B C, Fernandez I J, Norton S A. Mobilization of metals and phosphorus from intact forest soil cores by dissolved inorganic carbon. Environmental Monitoring and Assessment, 2010, 171(1/4): 93-110. |
| [24] | Ford C R, Wurzburger N, Hendrick R L, Teskey R O. Soil DIC uptake and fixation in Pinus taeda seedlings and its C contribution to plant tissues and ectomycorrhizal fungi. Tree Physiology, 2007, 27(3): 375-383. |
| [25] | Dawson J J C, Billett M F, Hope D, Palmer S M, Deacon C M. Sources and sinks of aquatic carbon in a peatland stream continuum. Biogeochemistry, 2004, 70(1): 71-92. |
| [26] | Fiedler S, Holl B S, Freibauer A, Stahr K, Drösler M, Schloter M, Jungkunst H F. Particulate organic carbon (POC) in relation to other pore water carbon fractions in drained and rewetted fens in Southern Germany. Biogeosciences, 2008, 5(6): 1615-1623. |
| [27] | Chow A T, Guo F M, Gao S D, Breuer R, Dahlgren R A. Filter pore size selection for characterizing dissolved organic carbon and trihalomethane precursors from soils. Water Research, 2005, 39(7): 1255-1264. |
| [28] | Michalzik B, Stadler B. Importance of canopy herbivores to dissolved and particulate organic matter fluxes to the forest floor. Geoderma, 2005, 127(3/4): 227-236. |
| [29] | Le Mellec A, Meesenburg H, Michalzik B. The importance of canopy-derived dissolved and particulate organic matter (DOM and POM)-comparing throughfall solutionfrom broadleaved and coniferous forests. Annals of Forest Science, 2010, 67(4): 411-411. |
| [30] | Schmidt M W I, Noack A G. Black carbon in soils and sediments: analysis, distribution, implications, and current challenges. Global Biogeochemical Cycles, 2000, 14(3): 777-793. |
| [31] | Auclair A N D, Carter T B. Forest wildfires as a recent source of CO2 at northern latitudes. Canadian Journal of Forest Research, 1993, 23(8): 1528-1536. |
| [32] | Kuhlbusch T A J. Black carbon and the carbon cycle. Science, 1998, 280(5371): 1903-1904. |
| [33] | Kerner M, Hohenberg H, Ertl S, Reckermann M, Spitzy A. Self-organization of dissolved organic matter to micelle-like microparticles in river water. Nature, 2003, 422(6928): 150-154. |
| [34] | Carvalho A, Pio C, Santos C, Alves C. Particulate carbon in the atmosphere of a Finnish forest and a German anthropogenically influenced grassland. Atmospheric Research, 2006, 80(2/3): 133-150. |
| [35] | Kuhlbusch T, Crutzen P. Toward a global estimate of black carbon in residues of vegetation fires representing a sink of atmospheric CO2 and a source of O2. Global Biogeochemical Cycles, 1995, 9(4): 491-501. |
| [36] | Wang C Y, Zhou J B, Xia Z M, Chen X L. Soluble organic carbon in plant litters on Loess Plateau: content and biodegradability. Chinese Journal of Applied Ecology, 2010, 21(12): 3001-3006. |
| [37] | Leff J W, Wieder W R, Taylor P G, Townsend A R, Nemergut D R, Grandy A S, Cleveland C C. Experimental litterfall manipulation drives large and rapid changes in soil carbon cycling in a wet tropical forest. Global Change Biology, 2012, 18(9): 2969-2979. |
| [38] | Michalzik B, Kalbitz K, Park J H, Solinger S, Matzner E. Fluxes and concentrations of dissolved organic carbon and nitrogen-a synthesis for temperate forests. Biogeochemistry, 2001, 52(2): 173-205. |
| [39] | Matamala R, Gonzàlez-Meler M A, Jastrow J D, Norby R J, Schlesinger W H. Impacts of fine root turnover on forest NPP and soil C sequestration potential. Science, 2003, 302(5649): 1385-1387. |
| [40] | Fan P P, Guo D L. Slow decomposition of lower order roots: a key mechanism of root carbon and nutrient retention in the soil. Oecologia, 2010, 163(2): 509-515. |
| [41] | Bond-Lamberty B, Wang C, Gower S T. A global relationship between the heterotrophic and autotrophic components of soil respiration? Global Change Biology, 2004, 10(10): 1756-1766. |
| [42] | Wang C K, Yang J Y. Rhizospheric and heterotrophic components of soil respiration in six Chinese temperate forests. Global Change Biology, 2007, 13(1): 123-131. |
| [43] | McDowell W H, Likens G E. Origin, composition, and flux of dissolved organic carbon in the Hubbard Brook Valley. Ecological Monographs, 1988, 58(3): 177-195. |
| [44] | Schmidt B H M, Wang C P, Chang S C, Matzner E. High precipitation causes large fluxes of dissolved organic carbon and nitrogen in a subtropical montane Chamaecyparis forest in Taiwan. Biogeochemistry, 2010, 101(1/3): 243-256. |
| [45] | Boissier J M, Fontvielle D. Biodegradable dissolved organic-carbon in seepage waters from two forest soils. Soil Biology & Biochemistry, 1993, 25(9): 1257-1261. |
| [46] | Lutz B D, Bernhardt E S, Roberts B J, Mulholand P J. Examining the coupling of carbon and nitrogen cycles in Appalachian streams: the role of dissolved organic nitrogen. Ecology, 2011, 92(3): 720-732. |
| [47] | Couture S, Houle D, Gagnon C. Increases of dissolved organic carbon in temperate and boreal lakes in Quebec, Canada. Environmental Science and Pollution Research, 2011, 19(2): 361-371. |
| [48] | Freeman C, Fenner N, Ostle N J, Kang H, Dowrick D J, Reynolds B, Lock M A, Sleep D, Hughes S, Hudson J. Export of dissolved organic carbon from peatlands under elevated carbon dioxide levels. Nature, 2004, 430(6996): 195-198. |
| [49] | Evans C D, Monteith D T, Cooper D M. Long-term increases in surface water dissolved organic carbon: observations, possible causes and environmental impacts. Environmental Pollution, 2005, 137(1): 55-71. |
| [50] | Badri D V, Vivanco J M. Regulation and function of root exudates. Plant, Cell & Environment, 2009, 32(6): 666-681. |
| [51] | Wang Q K, Wang S L. Soil organic matter under different forest types in Southern China. Geoderma, 2007, 142(3/4): 349-356. |
| [52] | Christ M J, David M B. Temperature and moisture effects on the production of dissolved organic carbon in a Spodosol. Soil Biology & Biochemistry, 1996, 28(9): 1191-1199. |
| [53] | Evans C D, Jones T G, Burden A, Ostle N, Zieliński P, Cooper M D A, Peacock M, Clark J M, Oulehle F, Cooper D, Freeman C. Acidity controls on dissolved organic carbon mobility in organic soils. Global Change Biology, 2012, 18(11): 3317-3331. |
| [54] | Bader M K F, Körner C. No overall stimulation of soil respiration under mature deciduous forest trees after 7 years of CO2 enrichment. Global Change Biology, 2010, 16(10): 2830-2843. |
| [55] | Ju W M, Chen J M, Black T A, Barr A G, Mccaughey H, Roulet N T. Hydrological effects on carbon cycles of Canada's forests and wetlands. Tellus Series B, 2006, 58(1): 16-30. |
| [56] | Buckingham S, Tipping E, Hamilton-Taylor J. Concentrations and fluxes of dissolved organic carbon in UK topsoils. Science of the Total Environment, 2008, 407(1): 460-470. |
| [57] | Hopkinson C S Jr, Vallino J J. Efficient export of carbon to the deep ocean through dissolved organic matter. Nature, 2005, 433(7022): 142-145. |
| [58] | Brookshire E N J, Valett H M, Thomas S A, Webster J R. Coupled cycling of dissolved organic nitrogen and carbon in a forest stream. Ecology, 2005, 86(9): 2487-2496. |
| [59] | Kindler R, Siemens J, Kaiser K, Walmsley David C, Bernhofer C, Buchmann N, Cellier P, Eugster W, Gleixner G, Grünwald T, Heim A, Ibrom A, Jones S K, Jones M, Klumpp K, Kutsch W, Larsen K S, Lehuger S, Loubet B, Mckenzie R, Moors E, Osborne B, Pilegaard K, Rebmann C, Saunders M, Schmidt M W I, Schrumpf M, Seyfferth J, Skiba U, Soussana J F, Sutton M A, Tefs C, Vowinckel B, Zeeman M J, Kaupenjohann M. Dissolved carbon leaching from soil is a crucial component of the net ecosystem carbon balance. Global Change Biology, 2011, 17(2): 1167-1185. |
| [60] | Aitkenhead J A, McDowell W H. Soil C: N ratio as a predictor of annual riverine DOC flux at local and global scales. Global Biogeochemical Cycles, 2000, 14(1): 127-138. |
| [61] | Bernhardt E S, Likens G E. Dissolved organic carbon enrichment alters nitrogen dynamics in a forest stream. Ecology, 2002, 83(6): 1689-1700. |
| [62] | Brunet F, Potot C, Probst A, Probst J L. Stable carbon isotope evidence for nitrogenous fertilizer impact on carbonate weathering in a small agricultural watershed. Rapid Communications in Mass Spectrometry, 2011, 25(19): 2682-2690. |
| [63] | Yang Y S, Guo J F, Chen G S, Chen Y X, Yu Z Y, Dong B, Liu D X. Origin, property and flux of dissolved organic matter in forest ecosystems. Acta Ecologica Sinica, 2003, 23(3): 547-558. |
| [64] | Clark J M, Ashley D, Wagner M, Chapman P J, Lane S N, Evans C D, Heathwaite A L. Increased temperature sensitivity of net DOC production from ombrotrophic peat due to water table draw-down. Global Change Biology, 2009, 15(4): 794-807. |
| [65] | Clark J M, Chapman P J, Heathwaite A L, Adamson J K. Suppression of dissolved organic carbon by sulfate induced acidification during simulated droughts. Environmental Science & Technology, 2006, 40(6): 1776-1783. |
| [66] | Henry H A L. Soil freeze-thaw cycle experiments: trends, methodological weaknesses and suggested improvements. Soil Biology and Biochemistry, 2007, 39(5): 977-986. |
| [67] | Hentschel K, Borken W, Matzner E. Repeated freeze-thaw events affect leaching losses of nitrogen and dissolved organic matter in a forest soil. Journal of Plant Nutrition and Soil Science, 2008, 171(5): 699-706. |
| [68] | Schmitt A, Glaser B, Borken W, Matzner E. Repeated freeze-thaw cycles changed organic matter quality in a temperate forest soil. Journal of Plant Nutrition and Soil Science, 2008, 171(5): 707-718. |
| [69] | Ågren A, Haei M, Köhler S, Bishop K, Laudon H. Long cold winters give higher stream water dissolved organic carbon (DOC) concentrations during snowmelt. Biogeosciences Discussions, 2010, 7(3): 4857-4886. |
| [70] | Monson R K, Lipson D L, Burns S P, Turnipseed A A, Delany A C, Williams M W, Schmidt S K. Winter forest soil respiration controlled by climate and microbial community composition. Nature, 2006, 439(7077): 711-714. |
| [71] | Feng X J, Nielsen L L, Simpson M J. Responses of soil organic matter and microorganisms to freeze-thaw cycles. Soil Biology & Biochemistry, 2007, 39(8): 2027-2037. |
| [72] | Wang Q K, Wang S L, Feng Z W. A study on dissolved organic carbon and nitrogen nutrients under Chinese fir plantation: relationships with soil nutrients. Acta Ecologica Sinica, 2005, 25(6): 1299-1305. |
| [73] | Clair T A, Arp P, Moore T R, Dalva M, Meng F R. Gaseous carbon dioxide and methane, as well as dissolved organic carbon losses from a small temperate wetland under a changing climate. Environmental Pollution, 2002, 116(Suppl 1): S143-S148. |
| [74] | Fiedler S, Holl B S, Jungkunst H F. Discovering the importance of lateral CO2 transport from a temperate spruce forest. Science of the Total Environment, 2006, 368(2/3): 909-915. |
| [75] | Jiang Y M, Chen C L, Xu Z H, Liu W Q, Ouyang J, Wang F. Soil soluble organic matter, microbial biomass, and enzyme activities in forest plantations in degraded red soil region of Jiangxi Province, China. Chinese Journal of Applied Ecology, 2010, 21(9): 2273-2278. |
| [76] | Lu X, Gilliam F, Yu G, Li L, Mao Q, Chen H, Mo J. Long-term nitrogen addition decreases carbon leaching in nitrogen-rich forest ecosystems. Biogeosciences Discussions, 2013, 10(1): 1451-1481. |
| [77] | Andrews D M, Lin H, Zhu Q, Jin L X, Brantley S L. Hot spots and hot moments of dissolved organic carbon export and soil organic carbon storage in the Shale Hills catchment. Vadose Zone Journal, 2011, 10(3): 943-954. |
| [78] | Manzoni S, Trofymow J A, Jackson R B, Porporato A. Stoichiometric controls on carbon, nitrogen, and phosphorus dynamics in decomposing litter. Ecological Monographs, 2010, 80(1): 89-106. |
| [36] | 王春阳, 周建斌, 夏志敏, 陈兴丽. 黄土高原区不同植物凋落物可溶性有机碳含量及其降解. 应用生态学报, 2010, 21(12): 3001-3006. |
| [63] | 杨玉盛, 郭剑芬, 陈光水, 陈银秀, 于占源, 董彬, 刘东霞. 森林生态系统DOM的来源、特性及流动. 生态学报, 2003, 23(3): 547-558. |
| [72] | 王清奎, 汪思龙, 冯宗炜. 杉木人工林土壤可溶性有机质及其与土壤养分的关系. 生态学报, 2005, 25(6): 1299-1305. |
| [75] | 江玉梅, 陈成龙, 徐志红, 刘苑秋, 欧阳菁, 王芳. 退化红壤区人工林土壤的可溶性有机物, 微生物生物量和酶活性. 应用生态学报, 2010, 21(9): 2273-2278. |