 2014, Vol. 34
2014, Vol. 34文章信息
- 冯永玖, 陈新军, 杨晓明, 高峰
- FENG Yongjiu, CHEN Xinjun, YANG Xiaoming, GAO Feng
- 基于遗传算法的渔情预报HSI建模与智能优化
- HSI modeling and intelligent optimization for fishing ground forecasts using a genetic algorithm
- 生态学报, 2014, 34(15): 4333-4346
- Acta Ecologica Sinica, 2014, 34(15): 4333-4346
- http://dx.doi.org/10.5846/stxb201212171813
-
文章历史
- 收稿日期:2012-12-17
- 修订日期:2014-3-3






2. 大洋渔业资源可持续开发省部共建教育部重点实验室, 上海 201306;
3. 国家远洋渔业工程技术研究中心, 上海 201306;
4. 远洋渔业协同创新中心, 上海 201306
2. The Key Laboratory of Sustainable Exploitation of Oceanic Fisheries Resources (Shanghai Ocean University), Ministry of Education, Shanghai 201306, China;
3. National Distant-water Fisheries Engineering Research Center (Shanghai Ocean University), Shanghai 201306, China;
4. Collaborative Innovation Center for Distant-water Fisheries, Shanghai 201306, China
栖息地适宜性指数(HSI)模型最早由美国科学家于20世纪80年代提出,用来模拟生物体对周围栖息环境要素的反应[1]。目前已广泛应用于物种分布与管理等领域[2],并逐渐在海洋渔场的分析与预测中得到了应用[3, 4, 5],如印度洋大眼金枪鱼[6, 7, 8]、大西洋及太平洋的鱿鱼[9, 10, 11]、以及鲐鱼、秋刀鱼等[12]。从研究方法来看,主要有权重求和法、几何平均法[6, 7]、分位数回归方法[13]和主成分分析法[14]等。HSI的研究不仅是为了从理论上认知渔场分布与海洋环境要素的关系,也是为了向捕捞生产和渔业资源管理者提供信息参考。
海洋环境要素与中心渔场之间存在动态交互关系并构成一个复杂的系统,环境要素之间通常存在一定的相关性。经典数理统计方法在构建HSI模型时,由于无法消除环境要素固有的多重相关性,分析预报的精度受到极大的限制。因此,如何建立高精度的渔情预报模型,降低预报的不确定性,便成为了鱼类HSI建模需要解决的关键科学问题。
从最小化中心渔场预测值与实测值之间的差异的角度出发,利用人工智能方法进行自动优化,则可望提高渔情预报的精度和降低预报的不确定性。作为一种著名的组合优化算法,遗传算法(GA)常用于求解复杂函数的极值[15]。遗传算法源自对达尔文生物进化论的模拟计算,是一种通过模拟自然进化过程搜索最优解的方法,目前已成为一种成熟的进化智能算法[16]。近20年来,遗传算法在众多领域得到了广泛应用,例如在复杂地理系统的建模中遗传算法被用于建立非线性元胞自动机模型[17],与之类似的组合优化算法粒子群优化算法和模拟退火算法也得到了较好的应用[18, 19]。鉴于此,本研究基于遗传算法构建一种能够获取和优化中心渔场HSI参数的智能建模框架GeneHSI,并利用模拟数据进行该框架的执行测试。同时,验证GeneHSI模型框架在一般优化策略、不等式、等式和上下界限制条件下对渔业模拟数据的建模优化,并测试了GeneHSI模型框架对于不同样本量数据的优化解算能力。该基于遗传算法的GeneHSI框架将有助于实现渔情预报HSI的优化建模,并可望引导智能方法在渔业HSI建模中的应用。
1 GeneHSI模型构建 1.1 遗传算法简介遗传算法(GA)是一种基于生物自然选择与遗传机理的随机搜索算法。与传统搜索算法不同,GA从一组随机产生的“种群” (初始可行解)开始搜索过程[15]。待解问题的每一个参数即是一个“基因”,一组基因构成了问题的一个可行解,解是种群中的个体,称为“染色体”。在遗传算法中,染色体被编码为一串符号(二进制字符串),这些字符串在在后续迭代中根据遗传规则不断进化[16]。遗传算法在适应度函数的引导下进行迭代搜索,通过适应度值来判断染色体的优劣并进行优胜劣汰;在未达到算法终止条件时,通过选择、交叉和变异3个算子产生下一代染色体。在形成新一代过程中,根据适应度值的大小选择部分后代,淘汰部分较差后代同时保持种群大小为常数,并继续进行随机搜索和迭代,直到算法收敛从而得到最优解(最优染色体)[15, 16]。
1.2 HSI与遗传算法空间的映射关系以综合栖息地基本模型(如连乘模型、算术平均模型、几何平均模型、logistic回归模型等)为基础,以实际作业换算的HSI为基准[20],构建GA算法的适应度函数[15]。利用遗传算法自动寻找该适应度函数的最小值,其意义为GA获取的参数能够使模型预测值与实际作业换算的HSI值之间的差异达到最小化[16]。对应于最小适应度函数值的染色体,就是一组通过GA优化所得的栖息地指数模型的参数,而对应的“基因”则为各海洋渔业环境要素的权重。由这样一组染色体构建的HSI模型,即为GA优化的渔场预报模型。
1.3 GeneHSI模型中GA的适应度函数与优化策略(1)染色体编码与种群初始化 染色体即是一组参数的可行解,采用浮点数编码方法,定义染色体的长度为有效解的变量长度,定义染色体编码为a={a0,a1,…,aj,…,am},aj是HSI模型参数,即GA算法中的基因,m表示基因个数。
(2)适应度函数 适应度函数是渔场预报建模向遗传算法映射的核心,只有构建适应度函数才能够理解遗传算法的本质,也才能够进行遗传算法的渔情预报HSI建模。基于我们前期对元胞自动机、模拟退火算法和粒子群算法的大量研究[16, 17, 18, 19],构建了渔情预报HSI建模的适应度函数。在遗传算法中,利用适应度函数F(a)评价染色体(可行解)的优劣,规定F(a)的值越小则对应的染色体越优:
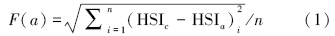
式中,HSIc表示通过海洋环境因素计算所得的渔场概率,即HSIc=1/[1+exp(-(a0+a1x1+…+amxm))],HSIa表示通过商业捕捞数据计算而得的渔场概率,n表示样本点的数量。其中,HSIc的表达式是一种logistic回归方程式,虽然目前为止这种表达形式在渔业栖息地研究中并未发现,但是logistic回归在野生动物的栖息地建模中广泛使用,一些经典论文的引用频次更是达到数百次之多,这表明logistic回归在动物栖息地建模中理论上是成立的[21, 22]。
国际上利用logistic回归进行野生动物栖息地建模的经典案例很多,包括格雷厄姆山红松鼠、高温蚱蜢和布什蟋蟀、高海拔稀有物种以及麂和喜马拉雅斑羚等[22, 23, 24, 25]。海洋鱼类是典型的水生野生动物,其栖息地模式与文献中野生动物非常接近,从生态学上可以判定logistic回归符合海洋鱼类。在一项关于鱼类种群判别的研究中,认为logistic回归比判别分析的效果要好,这直接证明了logistic回归可以用于渔业资源与渔场学领域[26]。在此将logistic回归方程作为核心构建适应度函数,但是在GeneHSI建模中并不需要真正执行logistic回归,只需要借用这种在理论上成立的方程式,通过遗传算法自动获取HSI参数从而完成HSI的建模。
(3)种群的选择、交叉与变异 确定适当的选择算子、交叉算子、变异算子以及算法终止条件,作为GA算法的优化策略[15, 16]。
(4)最优染色体的获取 最后获取的最优染色体即为需要确定的权重参数[15, 16],用于建立基于遗传算法的综合HSI模型框架GeneHSI。
基于遗传算法的基本理论以及从渔场预报向遗传算法的映射关系,构建用于渔情预报HSI参数获取与优化的建模框架GeneHSI(图 1)。GeneHSI模型框架通过Matlab“Optimization Tool”中的GA工具进行构建。由于该GA工具主要针对连续函数,而渔业HSI的适应度函数却是一种遍历所有样本点的离散函数,因此需要将该离散函数编写成为Matlab代码供GA工具调用和执行[16]。

|
| 图 1 遗传算法用于渔情预报HSI参数获取与优化的建模框架 Fig.1 A general framework for retrieving and optimizing fishing forecast HSI parameters based on generic algorithm |
图 1表明GeneHSI由3个部分组成:(1)待解问题构建,即HSI模型向遗传算法空间的映射,通过海洋环境因子、商业捕捞数据、以及基于logistic回归的渔场概率方程构建遗传算法的适应度函数;(2)GA初始化,即确定GA产生初始HSI参数的方案和GA初始种群数量等;(3)GA优化策略,即确定适应度函数优化过程中的迭代方法,包括可行解判断方法、新解产生的选择、交叉和变异算子以及算法终止规则等。其中,选择概率Ps一般通过比例选择和保存策略等方法自动确定,交叉概率Pc一般选取0.4—0.99之间的值,而变异概率Pm一般选取0.0001—0.1之间的值[15]。模型终止准则包括以下几个方面[15, 16]:1)GeneHSI最大计算时间(与计算机硬件关系较大),超出设定的时间则模型终止;2)最大迭代次数,达到该迭代次数则模型终止;3)适应度函数极限值,当达到极限值则模型终止;4)两次计算所得的适应度函数值的差值(计算精度),当该差值小于某阈值则模型终止。
2 GeneHSI框架的模拟数据应用与分析 2.1 渔业模拟数据为验证该GeneHSI模型框架的正确性和有效性,更进一步探测其使用和控制方法,使用模拟数据进行测试。在模拟数据中,假设与渔场概率相关的海洋环境因素有温度、盐度、叶绿素、溶解氧、温差以及海面高度距平均值等。由于各种海洋环境因素的值域范围不一致,为了能够在同一个方程式中进行换算,将其进行归一化处理,所有环境因子的值域范围归一化到[0,1]之间。在实际操作中,利用EXCEL的随机数发生器RAND()产生模拟数据,RAND()随机产生的数值均在[0—1]之间(表 1)。根据研究要求,分别产生了50、100、1000、5000和10000等5种样本数据,用于检验GeneHSI模型的有效性以及模型对样本量的响应。
|
数据项
No. of samples | 海表温度
Sea surface temperature (SST) | 盐度
Sea salinity (SS) | 叶绿素
Chlorophyll-a (CHA) | 溶解氧
Dissolved oxygen (DO) | 温差
Temperature difference (TD) | 海面高度
距平均值 Sea surface height anomaly (SSHA) | 渔场概率
Fishing ground probability (FGP) |
| 1 | 0.3003 | 0.2628 | 0.8899 | 0.2849 | 0.4754 | 0.8627 | 0.1919 |
| 2 | 0.6112 | 0.7253 | 0.5983 | 0.4180 | 0.9300 | 0.5017 | 0.7640 |
| 3 | 0.4074 | 0.8635 | 0.1323 | 0.5995 | 0.4017 | 0.7001 | 0.7629 |
| 4 | 0.2157 | 0.2779 | 0.2720 | 0.5612 | 0.2196 | 0.7543 | 0.6644 |
| 5 | 0.1506 | 0.4958 | 0.4862 | 0.3367 | 0.7677 | 0.9388 | 0.9665 |
| 6 | 0.7711 | 0.4506 | 0.4589 | 0.7334 | 0.1176 | 0.0071 | 0.9236 |
| 7 | 0.0386 | 0.6371 | 0.9703 | 0.0621 | 0.6235 | 0.6795 | 0.0886 |
| 8 | 0.1304 | 0.9272 | 0.6833 | 0.6298 | 0.6442 | 0.8488 | 0.8426 |
| 9 | 0.5302 | 0.1758 | 0.9313 | 0.2705 | 0.7852 | 0.8672 | 0.4537 |
| 10 | 0.5512 | 0.6660 | 0.3100 | 0.8085 | 0.3702 | 0.2512 | 0.0952 |
| … | … | … | … | … | … | … | … |
在GeneHSI模型执行之前,首先必须确定模型的优化策略,即确定遗传算子与终止准则。本文使用轮盘赌法自动确定选择概率Ps,选取交叉概率Pc为0.6,而变异概率Pm为0.1。由于模型计算时间与适应度极值的影响较小,本研究设置模型终止准则为最大迭代次数500次和最优适应度函数差值(计算精度)为1e-6,达到其中任何一个条件则模型终止。在GeneHSI模型实际执行中,均由于计算精度达到1e-6而终止模型运行。根据GeneHSI模型以及模型优化策略,首先利用50个模拟数据样本点建立了适应度函数,从而获取HSI的参数。遗传算法在获取HSI参数的过程中,受到适应度函数的引导并进行优化,随着优化的推进适应度函数值不断降低,最终达到最小值(图 2)。该最小值即为在最终获取的HSI参数下,预测渔场概率与真实渔场概率之间差异的最小值。

|
| 图 2 一般优化策略下利用遗传算法获取与优化HSI参数的收敛过程 Fig.2 Fitness track and scores of the corresponding fitness of GeneHSI model under a general optimization strategy |
遗传算法对HSI参数进行计算与优化的适应度收敛过程。最佳适应度值收敛到0.250918,平均适应度值收敛到0.27452,根据适应度函数的构造理论,适应度值越小表明HSI参数导致的预测预报误差越小。最佳适应度值和平均适应度值均随迭代而减小,最佳适应度值在第16次迭代时收敛,但平均适应度值上下波动,未呈现收敛趋势(图 2)。直到迭代结束,最佳、最差(Worst)与平均适应度值之间的差异仍较大,表明GeneHSI在没有限制参数的条件下收敛性较差,结果并不理想(图 2)。因此,获取的HSI参数与真实情况存在一定的差异(表 2)。
|
常数项
Constant | 海表温度
SST | 盐度
SS | 叶绿素
CHA | 溶解氧
DO | 温差
TD | 海面高度距平均值
SSHA |
| -1.1925 | -0.8240 | 0.2412 | 1.8855 | 0.9745 | 1.0139 | -0.8059 |
由于一般优化策略下获取的HSI参数并不理想,因此需对HSI的参数范围进行限制,使GeneHSI在有限的范围内进行搜索,确保结果更加准确。遗 传算法中,不等式、等式和参数范围限制条件较常用,其中不等式和等式限制条件指的是多种海洋环境因子(温度、盐度、叶绿素、溶解氧、温差以及海面高度距平均值)符合的某种关系,而参数范围限制条件指的是HSI参数上下界范围。一般地,不等式条件可以表达为:

式中,A和b是待定的限制条件,若A为矩阵,则b为向量;若A为向量,则b为数值;x是海洋环境因子向量。在GeneHSI优化执行中,x是既定的样本数据,A和b的设置需经过理论探讨和经验分析加以确定,可以源于传统方法的计算结果、也可以源于对研究对象和研究区域的认知。
为了探测不等式限制条件对GeneHSI结果的影响,设A=[1 1 1 1 1 1 1]、b=5进行GeneHSI模型执行,优化过程和HSI参数结果分别如图 3和表 3。

|
| 图 3 不等式限制条件下利用遗传算法获取与优化HSI参数的收敛过程 Fig.3 Fitness track and scores of the corresponding fitness of GeneHSI model under linear inequalities |
|
常数项
Constant | 海表温度
SST | 盐度
SS | 叶绿素
CHA | 溶解氧
DO | 温差
TD | 海面高度距平均值
SSHA |
| -0.7064 | -1.3827 | -0.1444 | 1.0329 | 1.2298 | 0.7182 | -0.0905 |
图 3表明,在不等式条件下最佳适应度值收敛到0.256759,平均适应度值收敛到0.264893。在迭代30次之后,平均与最佳适应度的收敛曲线较为接近,呈现较为明显的收敛状态(图 3)。当适应度函数曲线开始收敛,最佳与平均适应度值开始接近,而最差适与平均适应度值之间的差异仍较大(图 3)。
2.3.2 等式限制条件与不等式限制条件类似,等式限制条件可以表达为:

式中,Aeq和beq是待定的限制条件,若Aeq为矩阵,则beq为向量;若Aeq为向量,则beq为数值;x是海洋环境因子向量。与不等式条件类似,x是既定的样 本数据,Aeq和beq通过理论和经验分析加以确定。为探测等式限制条件对GeneHSI结果的影响,设置Aeq=[0 1 1 1 1 1 1]、beq=3.5进行优化。根据上述等式限制条件进行GeneHSI模型执行,其优化过程和HSI参数结果分别如图 4和表 4。

|
| 图 4 等式限制条件下利用遗传算法获取与优化HSI参数的收敛过程 Fig.4 Fitness track and scores of the corresponding fitness of GeneHSI model under linear equalities |
|
常数项
Constant | 海表温度
SST | 盐度
SS | 叶绿素
CHA | 溶解氧
DO | 温差
TD | 海面高度距平均值
SSHA |
| -1.3584 | -0.4125 | 0.9702 | 0.1394 | 0.7752 | 1.7820 | 0.1041 |
等式限制条件下,最佳适应度值收敛到0.284878,平均适应度值收敛到0.284905。在迭代25次之后,平均与最佳适应度值完全重合,说明适应度函数曲线完全收敛,且效果较好。随着适应度函数开始收敛,最佳、最差和平均适应度值完全一致,表明GeneHSI优化效果非常理想(图 4)。
2.3.3 上下界限制除了不等式和等式限制条件,在GeneHSI算法中也常用到上下界限制限制条件。研究表明,传统HSI建模中获取的HSI参数值域均为0到1之间[10, 11, 12, 13],因此在GeneHS优化控制中可利用这个关键的信息对适应度函数进行引导。为此,设置HSI参数的上界为 [1 1 1 1 1 1 1],而下界为[0 0 0 0 0 0 0],优化过程如图 5。

|
| 图 5 上下界限制条件下利用遗传算法获取与优化HSI参数的收敛过程 Fig.5 Fitness track and scores of the corresponding fitness of GeneHSI model under parameters′ bounds |
从图 5可知,上下界限制条件下GeneHSI在迭代到34次时开始收敛,最佳适应度值收敛到0.252479,平均适应度值收敛到0.253145,收敛后最佳与平均适应度曲线重合较好。随着优化进程最差和最佳适应度值之间的范围不断缩小,开始收敛到最优适应度值,但是优化中最差适应度值仍有局部反弹迹象(图 5)。总体评估来看,GeneHSI优化的效果相对较好,最终获取的HSI参数如表 5。
|
常数项
Constant | 海表温度
SST | 盐度
SS | 叶绿素
CHA | 溶解氧
DO | 温差
TD | 海面高度距平均值
SSHA |
| 0.7356 | 0.8166 | 0.2455 | 0.9838 | 0.7269 | 0.9995 | 0.5973 |
为达到GeneHSI模型的最佳优化效果,通常同时使用不等式、等式和上下界3种限制条件,形成综合限制条件。图 6显示了3种限制条件同时使用的适应度函数优化过程,具体限制条件与前述设置保持一致。

|
| 图 6 综合限制条件下利用遗传算法获取与优化HSI参数的收敛过程 Fig.6 Fitness track and scores of the corresponding fitness of GeneHSI model under comprehensive constraints |
综合限制条件下GeneHSI模型在迭代到27次时开始收敛,最佳适应度值收敛到0.380855,平均适应度值收敛到0.380919,收敛曲线表明优化效果非常理想。随着优化进程最差和最佳适应度值之间的范围收缩较快且非常彻底,收敛后曲线未发现任何波动(图 6),说明综合限制条件下GeneHSI取得了较佳的优化效果,获取的HSI参数如表 6。
|
常数项
Constant | 海表温度
SST | 盐度
SS | 叶绿素
CHA | 溶解氧
DO | 温差
TD | 海面高度距平均值
SSHA |
| 0.5077 | 0.7074 | 0.2709 | 0.5500 | 0.5668 | 0.6124 | 0.1986 |
除了各种限制条件,样本量对GeneHSI建模同样具有重要的影响。在渔场预报分析中,样本是通过商业捕捞而产生的,因此对于研究区的特定鱼种,样本量并不是研究者主观决定的。但是对于一种智能建模框架而言,应该具有处理大样本的能力。因此,本文进一步分析GeneHSI模型对于各种样本量的处理能力,并分析优化过程与优化结果对于不同样本量的响应。为此,产生了100、1000、5000和10000四种样本量,样本产生的方法如3.1节所述。选择在综合限制条件下进行GeneHSI模型执行,具体限制参数和前述设置一致,最终模型优化过程如图 7。

|
| 图 7 不同样本量下利用遗传算法获取与优化HSI参数的收敛过程 Fig.7 Fitness tracks and scores of the corresponding fitness of GeneHSI model under different samples |
图 7表明各种样本量在GeneHSI模型中均得到了优化处理,其中100样本量收敛于第21次迭代,最佳适应度值0.283916,平均适应度值0.283916;1000样本量收敛于第16次迭代,最佳适应度值0.288048,平均适应度值0.300328;5000样本量收敛于第13次迭代,最佳适应度值0.310632,平均适应度值0.311132;10000样本量收敛于第11次迭代,最佳适应度值0.308201,平均适应度值0.309327。模型优化过程与收敛曲线显示,10000样本量收敛最快,收敛效果最好;1000和5000样本量收敛效果次之;而100样本量收敛的效果较晚,完全收敛出现在第30次迭代。4种样本量在收敛之后,最差与平均适应度值之间存在局部的波动,其中大样本量10000的波动最为严重,而小样本量100的波动最小。优化结果表明,GeneHSI模型具有处理各种样本量的能力,同时对于样本量的响应也不尽一致,各种样本量下GeneHSI获取的参数如表 7所示。
|
样本数
Number of samples | 常数项
Constant | 海表温度
SST | 盐度
SS | 叶绿素
CHA | 溶解氧
DO | 温差
TD | 海面高度距平均值
SSHA |
| 100 | 0.0413 | 0.0703 | 0.0134 | 0.0268 | 0.1669 | 0.0854 | 0.0010 |
| 1000 | 0.1316 | 0.0180 | 0.0273 | 0.0097 | 0.0631 | 0.1302 | 0.0512 |
| 5000 | 0.0605 | 0.1808 | 0.2788 | 0.0830 | 0.1148 | 0.0926 | 0.0229 |
| 10000 | 0.0999 | 0.1191 | 0.0142 | 0.0024 | 0.4149 | 0.0141 | 0.0524 |
遗传算法用于渔场渔情预报,其核心思想在于可以使计算渔场概率值与真实渔场概率值的累计误差达到最小化。研究表明,遗传算法本质上是一种具有智能特性的随机优化算法[15],能够行之有效地效优化鱼类HSI的建模并获取HSI参数。由于遗传算法的随机特性[15],在对它进行控制并引导算法有效执行的过程中,对建模者有较高的技术要求(如对Matlab工具的熟练程度和Matlab代码编写的能力),需要研究者对研究区域与研究对象有较高的熟悉程度[16, 17]。本研究所采用的数据是随机产生的模拟数据,模拟数据的作用和关键在于:(1)模拟数据比之真实数据,其数据中存在的规律性要差,因此如果GeneHSI能在该模拟数据上得到很好的应用,那么证明GeneHSI模型具有很强的普适性,当应用于规律性更强的渔业真实数据时可望得到更好的结果;(2)遗传需要较强的人工干预与良好的参数控制经验[16, 17],模拟数据有利于从多角度探讨GeneHSI算法的控制与使用方法。这些使用控制方法为真实渔业数据的建模应用奠定了技术基础。
3.2 几种限制条件下GeneHSI优化效果的比较与一般优化策略下的GeneHSI比较,不等式条件下最佳适应度值要大、而平均适应度值要低,表明整体优化结果稍好。但是不等式条件下的GeneHSI优化效果并未达到最佳状态,表明在不等式条件下虽然效果优于一般优化策略,因此仍需对算法进行深入的优化控制。与一般优化策略和不等式优化策略比较,等式限制条件下GeneHSI参数的获取效果更好,优化曲线收敛更加彻底。
与一般优化策略和不等式优化策略比较,上下界限制条件下GeneHSI模型取得了更好的收敛效果;但比之等式条件,上下界条件下优化曲线的收敛情况却较差,表现为收敛较晚且最佳适应度曲线与平均适应度曲线重合性较差,但是却获得了更小的最佳和平均适应度值。综合限制条件与几种单项优化限制条件比较,GeneHSI模型取得了更好的收敛效果,适应度函数的收敛更早、且最佳与平均适应度值重合得非常一致。
总体来看,在不同的限制条件下,遗传算法获取的HSI具有较大的差异,其中一般优化策略下获取的HSI参数最差,加以限制的不等式、等式和上下界条件下其优化过程显著地更加合理,因此获取的HSI参数也更准确。理论上,适应度函数值是渔场概率计算值与真实值的累计误差[15, 16, 17],但是函数优化收敛过程较好时,最终对应的适应度值并不是所有限制条件下可能得到的最低值。因此,评价HSI参数结果是否合理和准确,并不是以适应度值的高低为唯一标准的[18, 19];应当同时考虑到适应度函数的优化收敛过程,并结合对研究区渔场的专业知识进行判断。综合来看,对GeneHSI的优化过程加以限制,尤其是上下界等限制条件的使用将使HSI参数优化结果更加合理[8, 9, 10]。
3.3 GeneHSI处理样本数据的效率针对于渔场预报HSI建模,增加样本量并不会显著增加遗传算法需要迭代的次数,也不会增加遗传算法的实际解算时间。这与地理复杂系统中的建模有所不同,后者会显著增加遗传算法的迭代次数和解算时间[18, 19]。这可能由两个原因造成的:本文使用的海洋环境因子相对较少,或渔业HSI建模与经典地理系统建模比较数据复杂程度稍低,因此遗传算法解算起来效率更高。
在对GeneHSI有效控制的前提下,大样本量反而有可能使遗传算法更早地收敛于合理的适应度值[15]。研究表明,大样本量得到的适应度值一般会大于小样本量得到的适应度值[16, 17]。但是这种情况并不是绝对的,当用于建模的数据规律性较强时,大样本量反而能够得到更小的适应度值[15]。在真实渔情预报中,受到商业捕捞数据的限制,一般很少能够获取到大样本量数据[6, 7, 8, 9, 10, 11],这进一步表明了GeneHSI能够高效地对渔场预报进行优化处理。
3.4 GeneHSI与传统HSI模型的比较传统HSI模型在渔业科学中应用广泛,主要用于栖息地质量评估、资源量估算、分析选择渔场和渔情预报等,但各自有不同的特点。针对不同目的和研究区,各种模型的适用性和效果则不同。与传统HSI模型比较,GeneHSI模型存在诸多不同之处(表 8)。
|
模型
Models | 单变量
Single variable | 多变量
Multi-variable | 变量相关性
Correlation of variables | 复杂性-智能化
Complexity and intellectualization | 参数意义明确
Explicit meaning of parameters | 模型特点
Model features |
| 关于连乘法、最小/最大值法、算术/几何平均法和分位数回归法的“模型特点”参见文献[27] | ||||||
| 连乘法 | √ | × | 不考虑 | 低-低 | × | 计算结果保守,对零值敏感 |
| 最小值法 | √ | × | 不考虑 | 低-低 | × | 估计结果保守,决定于最小SI (suitability index) 因子 |
| 最大值法 | √ | × | 不考虑 | 低-低 | × | 估计结果乐观,决定于最大SI 因子 |
| 算术平均法 | √ | × | 不考虑 | 低-低 | × | 估计结果折中,不受SI 极值的影响,未考虑单因素SI的权重 |
| 几何平均法 | √ | × | 不考虑 | 低-低 | × | 估计结果折中,效果较算术平均法差,参数越少则效果越优,受SI 零值影响较大 |
| 分位数回归法 | √ | × | 不考虑 | 中-低 | × | 能提供多种不同分位数的SI估计结果,阐释因变量的整体分配,结合前述几种模型计算最终HSI值,估计结果的保守或乐观程度决定于HSI计算模型的选取 |
| 遗传算法 | √ | √ | 自动消除 | 高-高 | √ | 同时考虑多种海洋环境因素、对因素的权重进行自动赋权与智能优化,估计结果的保守或乐观程度决定于GA限制参数 |
研究表明,经典的相关和回归分析方法并不适合于生态学领域的相关因素关系研究[27]。因此,渔业HSI建模一般地先采用单因子构建SI函数,再通过几何平均法[6, 7]、分位数回归方法[13]和主成分分析法[14]等计算HSI值。与表 8中的连乘法等6种模型不同,GeneHSI模型通过适应度函数直接搜索、优化并获取海洋环境因素的权重,进而直接计算得到HSI值。传统渔业HSI的应用研究实际上将各种不同类型的因素进行等值赋权,虽然这些方法都是经过渔业专家知识而获得的,但显然带有主观判断的成分[27]。在等值赋权中,海洋环境因子的相关性通常被忽略,而这种相关性将对HSI的建模具有负面的影响。GeneHSI模型同时考虑海表温度、盐度、叶绿素、溶解氧、温差和海面高度距平均值等不同类型的海洋环境因素,在遗传优化中自动消除这些因素相关性的负面影响。这表明,GeneHSI模型是一个典型的多变量HSI模型,且HSI参数的含义就是海洋环境因素的权重,而传统HSI模型是对SI曲线的一种拟合,其参数不具备物理意义。
比较而言,传统HSI模型的复杂性较低,因此其建模难度和智能化程度较低[6, 7, 27];而GeneHSI模型复杂性和建模难度均较高,但其具有高度的智能化程度。GeneHSI模型估计结果的保守或乐观程度决定于GA限制参数,在应用方面较适合于栖息地质量评估、渔场分析和渔情预报等。
4 结论基于广泛应用于各领域的遗传算法,本文提出了一种渔情预报HSI参数获取与智能优化的方法框架GeneHSI,并对该模型框架的使用和参数控制进行了分析与讨论。本研究的目的在于提供一种基于遗传算法的渔业HSI建模框架和行之有效的智能建模方法与思路,因此侧重于模型建立的整体框架、模型建立的思路以及模型执行过程中的人为控制。
基于遗传算法的HSI建模有助于发展现有的渔情预报的理论和方法。但在以后的研究中,应将该GeneHSI模型框架应用到实际的渔情预报分析中,同时通过实际的应用与反馈来优化GeneHSI模型。
| [1] | U. S. Fish and Wildlife Service. Habitat Evaluation Procedures (HEP). Washington: U. S. Fish and Wildlife Service, 1980. |
| [2] | U. S. Fish and Wildlife Service. Standards for the development of habitat suitability index models[R]. U. S. Fish and Wildlife Service, 1981: 1-81. |
| [3] | Store R, Jokimäki J. A GIS-based multi-scale approach to habitat suitability modeling. Ecological Modelling, 2003, 169(1): 1-15. |
| [4] | Jin L R, Sun K P, He H S, Zhou Y F. Research advances in habitat suitability index model. Chinese Journal of Ecology, 2008, 27(5): 841-846. |
| [5] | Zielinski W J, Dunk J R, Yaeger J S, LaPlante D W. Developing and testing a landscape-scale habitat suitability model for fisher (Martes pennanti) in forests of interior northern California. Forest Ecology and Management, 2010, 260(9): 1579-1591. |
| [6] | Fan W, Cui X S, Shen X Q. Progress in fishing-ground analysis and fishing condition forecasting. Journal of Fisheries of China, 2005, 29(5): 706-710. |
| [7] | Chen X J, Feng B, Xu L X. A comparative study on habitat suitability index of bigeye tuna, Thunnus obesus in the Indian Ocean. Journal of Fishery Sciences of China, 2008, 15(2): 269-278. |
| [8] | Fan J T, Chen X J, Qian W G, Liu B L. Distribution of fishing ground of Thunnus Alalunnus and its relationship with sea suderace temperature in the waters around Vanuatu. Transactions of Oceanology and Limnology, 2011, (1): 71-78. |
| [9] | Song L M, Zhou Y G. Developing an integrated habitat index for bigeye tuna (Thunnus obesus) in the Indian Ocean based on longline fisheries data. Fisheries Research, 2010, 105(2): 63-74. |
| [10] | Chen F, Chen X J, Liu B L, Qian W G, Tian S Q. Relationship between fishing ground of Ommastrephes bartramⅡ and vertical temperature structure in the northwestern Pacific Ocean. Journal of Shanghai Ocean University, 2010, 19(4): 495-504. |
| [11] | Chen X, Tian S, Chen Y, Liu B. A modeling approach to identify optimal habitat and suitable fishing grounds for neon flying squid (Ommastrephes bartramⅡ) in the Northwest Pacific Ocean. Fishery Bulletin, 2010, 108(1): 1-14. |
| [12] | Tian S Q, Chen X J, Chen Y, Xu L X, Dai X J. Evaluating habitat suitability indices derived from CPUE and fishing effort data for Ommatrephes bratramⅡ in the northwestern Pacific Ocean. Fisheries Research, 2009, 95(2/3): 181-188. |
| [13] | Li G, Chen X J, Guan W J. Stock assessment and risk analysis of management strategies for Scomber japonicus in the East China Sea and Yellow Sea using a Bayesian approach. Journal of Fisheries of China, 2010, 34(5): 740-750. |
| [14] | Feng B, Tian S, Chen X J. The habitat suitability index of Illex argentinus by using quartile regression method in the Southwest Atlantic. Transactions of Oceanology and Limnology, 2010, (1): 15-22. |
| [15] | Hu Z M, Chen X J, Zhou Y Q, Qian W G, Liu B L. Forecasting fishing ground of Dosidicus gigas based on habitat suitability index off Peru. Acta Oceanologica Sinica, 2010, 32(5): 67-75. |
| [16] | Xuan G N, Cheng R W. Genetic Algorithm and the Engineering Optimization. Yu X J, Zhou G G, translate. Beijing: Press of Tsinghua University, 2004. |
| [17] | Feng Y J, Liu Y, Han Z. Land use simulation and landscape assessment by using genetic algorithm based on cellular automata under different sampling schemes. Chinese Journal of Applied Ecology, 2011, 22(4): 957-963. |
| [18] | Feng Y, Liu Y. An optimised cellular automata model based on adaptive genetic algorithm for urban growth simulation//Yeh A, Shi J W, Leung Y, Zhou C, eds. Advances in Spatial Data Handling and GIS, Lecture Notes in Geoinformation and Cartography. Berlin, Heidelberg: Springer-Verlag, 2012: 27-38. |
| [19] | Feng Y J, Liu Y. A heuristic cellular automata approach for modelling urban land-use change based on simulated annealing. International Journal of Geographical Information Science, 2013, 27(3): 449-466. |
| [20] | Feng Y J, Liu Y, Tong X H, Liu M L, Deng S S. Modeling dynamic urban growth using cellular automata and particle swarm optimization rules. Landscape and Urban Planning, 2011, 102(3): 188-196. |
| [21] | Tian S Q, Chen X J. Impacts of different calculating methods for nominal CPUE on CPUE standardization. Journal of Shanghai Ocean University, 2010, 19(2): 240-245. |
| [22] | Keating K A, Cherry S. Use and interpretation of logistic regression in habitat-selection studies. Journal of Wildlife Management, 2004, 68(4): 774-789. |
| [23] | Pereira J M C, Itami R M. GIS-based habitat modeling using logistic multiple regression: A study of the Mt. Graham Red Squirrel. Photogrammetric Engineering and Remote Sensing, 1991, 57(11): 1475-1486. |
| [24] | Hein S, Voss J, Poethke H J, Boris S. Habitat suitability models for the conservation of thermophilic grasshoppers and bush crickets-simple or complex? Journal of Insect Conservation, 2007, 11(3): 221-240. |
| [25] | Singh N J, Yoccoz N G, Bhatnagar Y V, Fox J L. Using habitat suitability models to sample rare species in high-altitude ecosystems: a case study with Tibetan argali. Biodiversity and Conservation, 2009, 18(11): 2893-2908. |
| [26] | Singh A, Kushwaha S P S. Refining logistic regression models for wildlife habitat suitability modeling: A case study with muntjak and goral in the Central Himalayas, India. Ecological Modelling, 2011, 222(8): 1354-1366. |
| [27] | Prager M H, Fabrizio M C. Comparison of logistic regression and discriminant analyses for stock identification of anadromous fish, with application to striped bass (Morone saxatilis) and American shad (Alosa sapidissima). Canadian Journal of Fisheries and Aquatic Sciences, 1990, 47(8): 1570-1577. |
| [28] | Gong C, Chen X J, Gao F, Guan W J, Lei L. Review on habitat suitability index in fishery science. Journal of Shanghai Ocean University, 2011, 20(2): 260-269. |
| [3] | 金龙如, 孙克萍, 贺红士, 周宇飞. 生境适宜度指数模型研究进展. 生态学杂志, 2008, 27(5): 841-846. |
| [5] | 樊伟, 崔雪森, 沈新强. 渔场渔情分析预报的研究及其进展. 水产学报, 2005, 29(5): 706-710. |
| [6] | 陈新军, 冯波, 许柳雄. 印度洋大眼金枪鱼栖息地指数研究及其比较. 中国水产科学, 2008, 15(2): 269-278. |
| [7] | 范江涛, 陈新军, 钱卫国, 刘必林. 瓦努阿图周边海域长鳍金枪鱼渔场分布及其与表温关系. 海洋湖沼通报, 2011, (1): 71-78. |
| [9] | 陈峰, 陈新军, 刘必林, 钱卫国, 田思泉. 西北太平洋柔鱼渔场与水温垂直结构关系. 上海海洋大学学报, 2010, 19(4): 495-504. |
| [12] | 李纲, 陈新军, 官文江. 基于贝叶斯方法的东、黄海鲐资源评估及管理策略风险分析. 水产学报, 2010, 34(5): 740-750. |
| [13] | 冯波, 田思泉, 陈新军. 基于分位数回归的西南太平洋阿根廷滑柔鱼栖息地模型研究. 海洋湖沼通报, 2010, (1): 15-22. |
| [14] | 胡振明, 陈新军, 周应祺, 钱卫国, 刘必林. 利用栖息地适宜指数分析秘鲁外海茎柔鱼渔场分布. 海洋学报, 2010, 32(5): 67-75. |
| [15] | 玄光男, 程润伟. 遗传算法与工程优化. 于歆杰, 周根贵, 译. 北京: 清华大学出版社, 2004. |
| [16] | 冯永玖, 刘艳, 韩震. 不同样本方案下遗传元胞自动机的土地利用模拟及景观评价. 应用生态学报, 2011, 22(4): 957-963. |
| [20] | 田思泉, 陈新军. 不同名义CPUE计算法对CPUE标准化的影响. 上海海洋大学学报, 2010, 19(2): 240-245. |
| [27] | 龚彩霞, 陈新军, 高峰, 官文江, 雷林. 栖息地适宜性指数在渔业科学中的应用进展. 上海海洋大学学报, 2011, 20(2): 260-269. |