 2014, Vol. 34
2014, Vol. 34文章信息
- 解婷婷, 苏培玺, 周紫鹃, 张海娜, 李善家
- XIE Tingting, SU Peixi, ZHOU Zijuan, ZHANG Haina, LI Shanjia
- 荒漠绿洲过渡带沙拐枣种群结构及动态特征
- Structure and dynamic characteristics of Calligonum mongolicum population in the desert-oasis ecotone
- 生态学报, 2014, 34(15): 4272-4279
- Acta Ecologica Sinica, 2014, 34(15): 4272-4279
- http://dx.doi.org/10.5846/stxb201212101776
-
文章历史
- 收稿日期:2012-12-10
- 网络出版日期:2014-3-3






植物种群结构是种群最基本的特征,也是种群重要的特征之一,它不仅能反映种群动态及其发展趋势,在一定程度上也反映了种群与环境间的相关关系以及它们在种群中的地位[1, 2]。其相关研究可以反映种群数量特征、存活状态及生境适合度,同时结合种群的生态需求、存活和死亡状况及繁育能力,也能对种群的过去干扰和未来趋势做出估计与预测[3, 4],还可以展现种群生物学特性对环境条件的适应结果,对于濒危植物的保护和管理具有重要意义。
荒漠绿洲过渡带指荒漠生态系统和绿洲生态系统之间的生态交错带,是对气候变化较为敏感的区域之一。近年来,荒漠绿洲过渡带的重要性已经受到越来越多的高度重视,其中,环境干扰与植物种群和群落动态反映了过渡带与气候变化相互作用的机制,是理解和预测植被对气候变化响应的基础[5]。在我国黑河中游荒漠绿洲带分布着较多天然生长的沙拐枣,具有生长快、易繁殖、耐旱、抗风蚀、耐沙埋的特点,是优良的固沙先锋种,同时也是荒漠绿洲过渡带的主要优势种之一,对维护荒漠绿洲过渡带生态系统的稳定性起到了至关重要的作用,目前,关于沙拐枣的研究主要集中在种子萌发[6]、形态特性[7]、水分生理特性[8, 9]、解剖结构与光合特性[10, 11]和种群的扩张[12]等方面,此外,谭勇[13]等对我国沙拐枣属天然群落物种多样性与土壤因子的关系也进行了研究,但对处于过渡带特殊环境背景下该种群的结构特征及数量动态方面的研究还未见报道,而这正是物种保护和种群恢复的基础,鉴于此,本文旨在通过对荒漠绿洲过渡带沙拐枣种群的调查,试图揭示现有沙拐枣种群的状态,分析种群的年龄结构特征与受干扰情况,及其种群动态与内在的生活史特征和外在的干扰状况之间的互动关系,探讨种群更新与维持的机制,以期为沙拐枣管理以及合理保护和增殖、种群恢复策略的制定提供参考。
1 研究区概况与研究方法 1.1 研究区概况研究区地处甘肃省河西走廊中部临泽县北部,距中国生态系统研究网络临泽内陆河流域综合研究站约5 km,地理坐标(39°19′—39°21′ N,100°02′ —100°21′ E)。平均海拔1370 m,年平均降水量为117 mm,降水多集中在7—9月份,年潜在蒸发量2390 mm,为降水量的20多倍;年平均气温7.6 ℃,最高气温39.1 ℃,最低-27 ℃,≥10 ℃的年积温为3088 ℃,属干旱荒漠气候类型,风向以西北风为主,年平均风速为3.2 m/s,最大风速21 m/s,>8级大风日数年平均为15 d,地带性土壤为灰棕漠土。地带性植被有沙拐枣、泡泡刺(Nitraria sphaerocarpa Maxim.)、红砂(Reaumuria soongorica Maxim.)、籽蒿( Artemisia sphaerocephala Krasch.)、盐生草(Halogeton glomeratus(Bieb.) C. A. Mey.)、雾冰藜(Bassia dasyphylla(Fish. Et Mey.)O. Kuntze)、细叶骆驼蓬(Peganum nigellastrum Bge.)和沙蓬(Agriophyllum squarrosum(Linn.) Moq.)。
1.2 研究方法 1.2.1 样地设置与调查方法在黑河中游临泽县北部沙质荒漠绿洲过渡带沙拐枣样带,从绿洲边缘开始,从南向北延伸,长1000 m。以绿洲边缘为起点,每隔120 m由近至远设置A1(距绿洲20 m)、A2、 A3、A4、A5、A6、A7、A8、A9共9个样地。样地大小为40 m×40 m,在每个样地中,再以20 m为间隔设置成4个20 m×20 m的样方,记录每种灌木的个体数、盖度、基径、高度、冠幅等指标;同时在每个20 m×20 m的样方内随机取3个1 m ×1 m的草本样方,调查草本种类、数量、盖度、高度、频度等指标。
1.2.2 径级划分虽然种群的龄级和径级有所不同,但同一环境下,同一树种的龄级和径级对环境的反应规律具有一致性[14],对于数量较少的濒危树种,在很难获取其年龄的情况下,大多数学者采用径级结构代替年龄级结构[15, 16],因而本文也采用径级结构代替年龄结构分析优势种群的结构动态特征。结合本次调查实际情况,由于大多数沙拐枣主干不明显,就以沙拐枣基径作为划分标准。根据基径(BD)的分布状况,将基径划分为10个等级:Ⅰ级(BD<0.5 cm)、Ⅱ级(0.5 cm≤BD<l.0 cm )、Ⅲ级(1.0 cm≤BD <1.5 cm)、Ⅳ级(1.5 cm≤BD<2.0 cm)、Ⅴ级(2.0 cm≤BD<2.5 cm)、Ⅵ级(2.5 cm≤BD<3.0 cm)、Ⅶ级(3.0 cm≤BD<3.5 cm)、Ⅷ级 (3.5 cm≤BD<4.0 cm)、Ⅸ级(4.0 cm≤BD<4.5 cm)和X级(BD≥4.5cm)。根据上述大小级划分标准,以大小级为横轴,以各大小级的株数为纵轴分别作优势种群大小级分布图。
1.2.3 种群动态数量化方法种群动态数量化方法采用文献[17]的方法,定量描述种群动态:
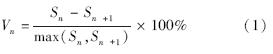
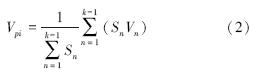
式中,Vn为种群从n到n+1级的个体数量变化;Vpi为整个种群结构的数量变化动态指数;Sn、Sn+1分别为第n和n+1级种群个体数;k为种群大小级数量.式(2)仅适用于不考虑未来外部环境干扰的种群结构动态的比较,当考虑未来的外部干扰时,则种群结构动态还与大小级数量(k)及各大小级个体数(S )两因素相关,因此将式(2)修正为:
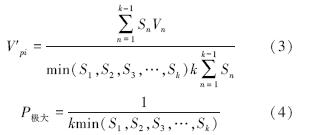
式中,Vn、Vpi 和V′pi分别反映种群个体数量的增长、衰退和稳定的动态关系.
1.2.4 种群静态生命表编制方法静态生命表又称特定时间生命表,多用于对长寿命的木本植物种群的统计研究,根据对不同年龄径级每年等量生长的假设,本文在沙拐枣的径级划分标准的基础上,综合样地数据对个体按0.5 cm 的级差编制静态生命表进行精细分析,详细编制方法参考文献[18]。
1.2.5 种群数量动态的时间序列分析本文采用时间序列分析的一次移动平均法[19, 20]对沙拐枣的种群龄级结构进行模拟和预测。
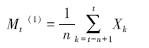
式中,n表示需要预测的未来时间年限,t为龄级,是近期n个观测值在t时刻的平均值,称为第n周期的移动平均,即表示未来n年时t龄级的种群大小,Xk为当前k龄级的种群大小。本文t分别取种群经历2、4、6和8龄级的时间后,对种群未来发展趋势进行预测。
2 结果与分析 2.1 沙拐枣种群分布现状及结构特征由图 1可以看出,荒漠绿洲过渡带沙拐枣种群龄级呈现基部极宽顶部狭窄的金字塔型,经统计,沙拐枣种群在Ⅰ—Ⅳ龄级个体数比较丰富,占整个种群个体数的89.98%,Ⅴ—Ⅶ龄级占8.4%,Ⅷ—Ⅹ龄级仅占1.62%,种群整体结构接近金字塔型,但沙拐枣种群中Ⅰ、Ⅱ龄级个体数目少于Ⅲ龄级的个体数目。

|
| 图 1 沙拐枣种群龄级结构 Fig. 1 The size structure of Calligonum mongolicum population |
根据种群动态量化方法分析沙拐枣种群相邻大小级个体变化,以对种群动态进行更加客观精确的评价。由表 1可以看出,沙拐枣种群相邻各级间个体数量变化动态指数为V1、V2小于0,说明Ⅰ、Ⅱ龄级个体数目少于Ⅲ龄级个体数目。在不考虑外界的环境干扰时,沙拐枣种群大小级结构的动态指数Vpi=0.2333>0,受随机干扰时的种群年龄结构动态指数Vpi′=0.0233>0,但趋于零,随机干扰风险极大值即种群结构对随机干扰的敏感性指数P=0.1,说明荒漠绿洲过渡带沙拐枣种群为增长型,但种群结构增长性低,趋于稳定型,且对外界干扰较敏感。
| 种群动态指数级 Dynamic index | 动态指数值 Dynamic index value/% |
| V1 | -3.33 |
| V2 | -3.85 |
| V3 | 32.05 |
| V4 | 69.81 |
| V5 | 65.62 |
| V6 | 18.18 |
| V7 | 44.44 |
| V8 | 20 |
| V9 | 75 |
| Vpi | 23.33 |
| Vpi′ | 2.33 |
由于静态生命表用特定时间段调查的种群所有个体编制而成,荒漠绿洲过渡带沙拐枣种群是天然种群,分析中利用异龄林种群年龄结构在空间上的差异研究特定时间断面上的种群动态,难免存在系统抽样误差产生与数学假设不符的现象,但仍能提供有用的生态学记录。对于这种情况,在编制静态生命表时通常会采用匀滑(Smooth out)技术,这里采用方程拟合方法进行匀滑处理。把大小级作为自变量,存活株数作为因变量,对其进行拟合,拟合方程为y=528.5e-0.5808x(R2=0.9394)。从静态生命表(表 2)可以看出,荒漠绿洲过渡带沙拐枣种群随年龄增加,存活数lx逐渐减少。个体的生命期望值ex在I、Ⅱ级时最大,随后逐渐降低。
存活曲线是借助于存活个体数量来描述特定年龄死亡率,它是通过把特定年龄组的个体数量相对作图而得到的。其绘制方法有两种,一是以存活量的对数值lnlx为纵坐标,以年龄为横坐标作图;另一种方法是用存活数量对年龄作图,但年龄用平均寿命期望的百分离差来表示。本文以存活量lx为纵坐标,以龄级为横坐标作图(图 2)。按Deevey 的划分,一般有3 种基本类型,Ⅰ型是凸曲线,属于该型的种群绝大多数都是能活到该物种年龄,早期死亡率较低,但当活到一定生理年龄时,短期内几乎全部死亡;Ⅱ型是直线,也称对角线型,属于该型的种群各年龄的死亡率基本相同;Ⅲ型是凹曲线,早期死亡率高,一旦活到某一年龄,死亡率就较低。由图 2可以看出,沙拐枣的存活曲线表明沙拐枣早期死亡数较高,曲线斜率较大,环境筛的选择强度大,只有5.4% 的幼树能穿过此筛进入Ⅵ龄级。Hett和Loucks 在检验估算的存活状况是符合Deevey-Ⅱ型曲线还是符合Deevey-Ⅲ型曲线时,采用两种数学模型进行检验,即指数方程式Nx=N 0e-bx 用以描述Deevey-Ⅱ型存活曲线,幂函数式N x=N0x-b描述Deevey-Ⅲ型存活曲线[21]。这里采用上述两种模型进行沙拐枣种群存活曲线类型的检验,经建立其相应模型得到:
N x = 1685.9e-0.56538x (R2= 0. 9983)
N x = 2207.9x-2.2372(R2= 0.9163)
由于指数模型的R2值(0.9983)大于幂函数模型的R2值(0.9163),因此,可以认为沙拐枣种群存活曲线更趋于Deevey-Ⅱ,呈对角线型,各年龄级种群有相近的死亡率。
| 龄级 x Age class | 基径 BD Interval/cm | Ax | ax | lx | dx | qx | Lx | Tx | ex | lnlx | Kx |
| x: 年龄级;Ax: 在x龄级的实际存活数;ax: 匀滑后的x龄级的存活数;lx: 在x龄级开始时标准化存活数(以1 000为基数),lx= ax/ a0×1000;dx: 从x到x+1龄级期间标准化死亡数,dx= lx- lx+1;qx: 从x到x+1龄级间隔期间的死亡率,qx= dx/lx;Lx: 从x到x+1龄级间隔期间平均存活的个体数,Lx.=( lx+lx+1)/2;Tx: 从x龄级到超过x龄级的个体总数,Tx = Lx+ Lx+1 + Lx+2+口,即Tx =∑Lx;ex: 进入 x龄级个体的生命期望,ex = Tx/ lx;Kx: 消失率(损失度),Kx= lnlx-lnlx+1 | |||||||||||
| Ⅰ | BD<0.5cm | 145 | 296 | 1000 | 443 | 0.44 | 779 | 1762 | 1.76 | 6.91 | 0.59 |
| Ⅱ | 0.5≤BD<l.0cm | 150 | 165 | 557 | 243 | 0.44 | 436 | 983 | 1.76 | 6.32 | 0.57 |
| Ⅲ | 1.0≤BD<l.5cm | 156 | 93 | 314 | 138 | 0.44 | 245 | 547 | 1.74 | 5.75 | 0.58 |
| Ⅳ | 1.5≤BD<2.0cm | 106 | 52 | 176 | 78 | 0.44 | 137 | 302 | 1.72 | 5.17 | 0.59 |
| V | 2.0≤BD<2.5cm | 32 | 29 | 98 | 44 | 0.45 | 76 | 165 | 1.68 | 4.58 | 0.59 |
| Ⅵ | 2.5≤BD<3.0cm | 11 | 16 | 54 | 24 | 0.44 | 42 | 89 | 1.65 | 3.99 | 0.59 |
| Ⅶ | 3.0≤BD<3.5cm | 9 | 9 | 30 | 13 | 0.43 | 24 | 47 | 1.57 | 3.40 | 0.57 |
| Ⅷ | 3.5≤BD<4.0cm | 5 | 5 | 17 | 7 | 0.41 | 14 | 23 | 1.35 | 2.83 | 0.53 |
| Ⅸ | 4.0≤BD<4.5cm | 4 | 3 | 10 | 3 | 0.30 | 9 | 9 | 0.90 | 2.30 | 0.35 |
| X | BD≥4.5cm | 1 | 2 | 7 | - | - | - | - | - | 1.95 | - |
沙拐枣种群死亡率(qx)和消失率(Kx)曲线变化趋势基本一致(图 2),沙拐枣种群的死亡率和消失率在Ⅸ龄级以前变化幅度不大,基本相近,这与存活曲线的分析结果相一致,只有在Ⅸ龄级个体向Ⅹ级阶段的发展过程中,死亡率和消失率显著降低,死亡率为0.3,消失率为0.35。

|
| 图 2 沙拐枣种群存活曲线和死亡率及消失率曲线 Fig. 2 Standard survival,mortality and vanish rate curve of Calligonum mongolicum population |
沙拐枣种群数量动态时间序列预测以沙拐枣种群各龄级株数为原始数据,按照一次平均推移法预测出各龄级在未来2、4、6、8个龄级时间后的个体数。可以看出(表 3),在未来2、4、6、8个龄级时间后,荒漠绿洲过渡带沙拐枣种群从Ⅳ龄级后的个体数均呈现小幅度的增加趋势,表明沙拐枣幼龄个体占总数的比例高,使得种群生长中后期的个体数得到一定的补充。而沙拐枣Ⅱ龄级在经历未来2个龄级时间后,个体数从原来的150株下降到148株,Ⅲ龄级在经历未来2个龄级时间后,个体数从原来的156株下降到153株,这一结果表明,由于沙拐枣幼苗的不足,小树数量将有所衰减,但幅度不大。可以推断,由于缺乏可更新的幼龄个体,如不采取适当的护林抚育措施,沙拐枣种群未来必然趋于衰退。
| 龄级 X Age class | 原始数据 Primary data | M(1)2 | M(1)4 | M(1)6 | M(1)8 |
| Ⅰ | 145 | ||||
| Ⅱ | 150 | 148 | |||
| Ⅲ | 156 | 153 | |||
| Ⅳ | 106 | 131 | 139 | ||
| V | 32 | 69 | 111 | ||
| Ⅵ | 11 | 22 | 76 | 100 | |
| Ⅶ | 9 | 10 | 40 | 77 | |
| Ⅷ | 5 | 7 | 14 | 53 | 77 |
| Ⅸ | 4 | 5 | 7 | 28 | 59 |
| X | 1 | 3 | 5 | 10 | 41 |
植物种群表现结构是种群发育过程中每个个体实现其增长机会的一种表达,也是对立地条件优劣及植物对环境适应性的反映[22]。比较不同种群的大小结构能反映植物与环境之间的适合度。本研究发现荒漠绿洲过渡带沙拐枣种群的年龄结构呈非典型金字塔型,种群呈现增长型,有向稳定型转化的趋势,这说明沙拐枣种群适应当地的生境条件,并能利用当地的环境条件实现自身生存扩展的最大化。但不容忽视的是,沙拐枣种群年龄结构Ⅰ、Ⅱ龄级数量少于Ⅲ龄级个体数量,说明沙拐枣幼龄级株数少,Ⅲ级株数较多,虽然种群总体呈现增长,但Ⅰ、Ⅱ龄级到Ⅲ龄级呈衰退趋势,也就是说,如果没有幼龄个体对中老龄个体的补充,沙拐枣种群整体的长期稳定性将难以维持,这与其他一些频危植物情况相似[23]。河西走廊荒漠绿洲过渡带沙质生境中沙拐枣主要以有性繁殖来保证幼苗的数量,同时沙拐枣无性繁殖也可以产生一定数量的幼龄个体[12],从而使得该物种具备数量相对丰富的幼龄个体,但由于幼龄个体竞争能力较弱,在其生长过程中最易受到动物采食和不利气候因素的影响,沙拐枣幼龄个体很难进入种群的更替层,即种群幼龄植株很难向中龄和成龄阶段转化,这在一定程度上限制了沙拐枣种群的自然更新,形成该种群更新和发展的瓶颈。
3.2 荒漠绿洲过渡带沙拐枣种群动态趋势生命表是判断植物种群发展趋势的重要指标之一,对植物种群的生命表和存活曲线的分析可以反映种群现实状况、展现植物种群与环境的竞争关系,对珍稀濒危植物的保护和利用具有重要意义[24]。本研究表明,荒漠绿洲过渡带沙拐枣种群的存活曲线为Deevey-Ⅱ,种群在Ⅸ龄级以前的死亡率基本相近,这说明沙拐枣种群各龄级树种的死亡率都较高,这与大多数研究发现幼龄阶段的种群死亡率最高的结果不相一致[23, 25, 26]。分析原因可能是与沙拐枣个体经受的环境阻力和个体的竞争力大小有关,沙拐枣种群Ⅰ—Ⅲ龄期个体数量较为丰富,且个体比较脆弱,环境阻力(包括土壤水分、养分等)使得其大量死亡,只有一部分进入到Ⅳ—Ⅶ龄期,此阶段可能由于大量的沙拐枣被人为樵采,从而导致死亡率较高,而Ⅷ—Ⅸ龄期可能由于随着年龄增长,种群的抗性及适应环境的能力随之减弱,对于沙拐枣种群来说其利用土壤养分、水分等条件生长的能力下降,使种群的存活率开始下降。从时间序列预测来看,在未来2、4、6、8个龄级时间后沙拐枣老龄个体逐渐增多,幼龄株数则有所降低,这说明由于缺乏可更新的幼龄个体,如不加强幼龄期的抚育管理,沙拐枣种群未来必然趋于衰退。
3.3 荒漠绿洲过渡带沙拐枣适应性保护和恢复对策对于同时具有两种繁殖方式的沙拐枣种群来说,促进其种子向幼苗转化率是种群恢复的关键环节,成年沙拐枣的结实量较大,保护好荒漠绿洲过渡带现有的成年个体,提高成年个体种子生产能力,促进种子的萌发和幼苗的更新,沙拐枣种群恢复潜力还是存在的,同时也要保证沙拐枣的无性繁殖个体的成活率,而且在沙拐枣种子丰产年,适时采集一些种子,进行撒播、飞播,提高天然条件下的种子发芽率,通过人工辅助进行种群的恢复。因此,适当的人工辅助恢复和现有植株和生境的保护是保持沙拐枣种群自然更新和进行种群恢复的关键。
| [1] | Wang Z F, An S Q, Zhu X L, Campell D G, Yang X B. Distribution pattern of tree populations in tropical forest and comparison of its study methods. Chinese Journal of Applied Ecology, 1998, 9(6): 575-580. |
| [2] | Li J Q. Forest Ecology. Beijing: Higher Education Press, 2006: 186-192. |
| [3] | Harper J L. Population Biology of Plants. London: Academic Press, 1977. |
| [4] | Zu Y G, Zhang W H, Yan X F. Conservation Biology of Adenophora iobophylla, an Endangered Plant. Beijing: Science Press, 1999. |
| [5] | Ma Z M, Pan B R. The Classification and distribution of the genus Calligonum L. in China. Acta Phytotaxonomica Sinica, 1986, 24(2): 98-107. |
| [6] | Ren J, Tao L. Effect of hydration-dehydration cycles on germination of seven Calligonum species. Journal of Arid Environments, 2003, 55(1): 111-122. |
| [7] | Singh G. Influence of soil moisture and nutrient gradient on growth and biomass production of Calligonum polygonoides in Indian desert affected by surface vegetation. Journal of Arid Environments, 2004, 56(3): 541-558. |
| [8] | Dhief A, Gorai M, Aschi-Smiti S, Neffati M. Comparative phenological and water potential patterns of three Calligonum species in the eastern great Erg of Tunisia. Flora, 2009, 204(8): 581-592. |
| [9] | Shan L S, Li Y, Zhao R F, Zhang X M. Effects of deficit irrigation on daily and seasonal variations of trunk sap flow and its growth in Calligonum arborescens. Journal of Arid Land, 2013, 5(2): 233-243. |
| [10] | Su P X, Zhao A F, Zhang L X, Du M W, Chen H S. Characteristic in photosynthesis, transpiration and water use efficiency of Haloxylon ammodendron and Calligonum mongolicum of desert species. Acta Botanica Boreali-Occidentalia Sinica, 2003, 23(1): 11-17. |
| [11] | Qiu Z J, Li Y, Chong P F. Effects of PEG stress on physiological characteristics of Calligonum mongolicum in different geographical provenance. Acta Prataculturae Sinica, 2011, 20(3): 108-114. |
| [12] | Zhuang Y L, Zhao W Z, Xie G X, Hu G L. The features of Calligonum mongolicum population and its expansion process in sandy habitat. Acta Ecologica Sinica, 2008, 28(4): 1399-1407. |
| [13] | Tan Y, Zhang Q, Pan B R, Kang X S. Coupling relationship between Calligonum L. community species diversity and soil factors in China. Arid Land Geography, 2008, 31(1): 88-96. |
| [14] | Frost I, Rydin H. Spatial pattern and size distribution of the animal-dispersed tree Quercus robur in two spruce-dominated forests. Ecoscience, 2000, 7(1): 38-44. |
| [15] | Ma K, Li G Y, Zhu C X, Xia G H. Population structure and distribution patterns of the rare and endangered Ardisia violacea (Myrsinaceae). Acta Ecologica Sinica, 2013, 33(2): 72-79. |
| [16] | Kang H J, Chen Z L, Liu P, Hao C Y, Wei F M. The population structure and distribution pattern of Emmenopterys henryi in Dapanshan natural reserve of Zhejiang Province. Acta Ecologica Sinica, 2007, 27(1): 389-396. |
| [17] | Chen X D. A study on the method of quantitative analysis for plant population and structural dynamics. Acta Ecologica Sinica, 1998, 18(2): 214-217. |
| [18] | Xie Z Q, Chen W L, Lu P, Hu D. The demography and age structure of the endangered plant population of Cathaya argyrophylla. Acta Ecologica Sinica, 1999, 19(4): 523-528. |
| [19] | Shen S K, Ma H Y, Wang Y H, Wang B Y, Shen G Z. The structure and dynamics of natural population of the endangered plant Euryodendron excelsum H. T. Chang. Acta Ecologica Sinica, 2008, 28(5): 2404-24l2. |
| [20] | Guo Q Q, Lu J, Luo D Q, Fang J P, Wang Z H. Ecological characteristics of Sabina saltuaria population at the timberline ecotone on the sunny slope of the Sejila Mountain. Journal of Northwest Forestry University, 2010, 25(2): 15-18. |
| [21] | Silvertown J W. Introduction to Plant Population Ecology. London: Longman Group United Kingdom, 1982. |
| [22] | Zhang J, Huang Y M, Jiao H J, Xian X D Ecophysiological characteristics of artemisia ordosica community succession in Mu Us sandy grassland. Journal of Desert Research, 2007, 27(6): 977-983. |
| [23] | Li X X, Wang Q C, Cui G F, Yang C D. Structure and dynamic characteristics of a wild population of endangered Abies fanjingshanensis. Acta Botanica Boreali-Occidentalia Sinica, 2011, 31(7): 1479-1486. |
| [24] | Wang Z H, Huang R F, Wang I H, Zhang G S. Analysis on life table of natural Sabina vulgaris population in Mu Us sandy land. Journal of Desert Research, 2009, 29(1): 118-124. |
| [25] | Wang D J, Wen G S, Tong Z K, Xu J Q, Wu J, Liu T X. Analysis to the static life table of Casuarina Equisetifolia populations on the big deer island in Zhejiang. Journal of Inner Mongolia Agricultural University: Natural Science Edition, 2011, 32(4): 145-150. |
| [26] | Ren Q S, Tang X L, Cui G F, Wang J S, Huang Y, Wei X H, Li Q L. Smith fir population structure and dynamics in the timberline ecotone of the Sejila Mountain, Tibet, China. Acta Ecologica Sinica, 2007, 27(7): 2669-2677. |
| [1] | 王峥峰, 安树青, 朱学雷, Campell D G, 杨小波. 热带森林乔木种群分布格局及其研究方法的比较. 应用生态学报, 1998, 9(6): 575-580. |
| [2] | 李俊清. 森林生态学. 北京: 高等教育出版社, 2006: 186-192. |
| [4] | 祖元刚, 张文辉, 阎秀峰. 濒危植物裂叶沙参保护生物学. 北京: 科学出版社, 1999. |
| [5] | 毛祖美, 潘伯荣. 我国沙拐枣属的分类与分布. 植物分类学报, 1986, 24(2): 98-107. |
| [10] | 苏培玺, 赵爱芬, 张立新, 杜明武, 陈怀顺. 荒漠植物梭梭和沙拐枣光合作用、蒸腾作用及水分利用效率特征. 西北植物学报, 2003, 23(1): 11-17. |
| [11] | 邱真静, 李毅, 种培芳. PEG胁迫对不同地理种源沙拐枣生理特性的影响. 草业学报, 2011, 20(3): 108-114. |
| [12] | 庄艳丽, 赵文智, 谢国勋, 胡光录. 沙地生境沙拐枣(Calligonum mongolicum)种群特征及其扩张. 生态学报, 2008, 28(4): 1399-1407. |
| [13] | 谭勇, 张强, 潘伯荣, 康晓珊. 我国沙拐枣属(Calligonum L.)天然群落物种多样性与土壤因子的耦合关系. 干旱区地理, 2008, 31(1): 88-96. |
| [16] | 康华靖, 陈子林, 刘鹏, 郝朝运, 韦福民. 大盘山自然保护区香果树种群结构与分布格局. 生态学报, 2007, 27(1): 389-396. |
| [17] | 陈晓德. 植物种群与群落结构动态量化分析方法研究. 生态学报, 1998, 18(2): 214-217. |
| [18] | 谢宗强, 陈伟烈, 路鹏, 胡东. 濒危植物银杉的种群统计与年龄结构. 生态学报, 1999, 19(4): 523-528. |
| [19] | 申仕康, 马海英, 王跃华, 王博轶, 申国柱. 濒危植物猪血木(Euryodendron excelsum H. T. Chang)自然种群结构及动态. 生态学报, 2008, 28(5): 2404-24l2. |
| [20] | 郭其强, 卢杰, 罗大庆, 方江平, 王贞红. 西藏色季拉山阳坡林线方枝柏种群结构特征研究. 西北林学院学报, 2010, 25(2): 15-18. |
| [22] | 张军, 黄永梅, 焦会景, 夏显东. 毛乌素沙地油蒿群落演替的生理生态学机制. 中国沙漠, 2007, 27(6): 977-983. |
| [23] | 李晓笑, 王清春, 崔国发, 杨传东. 濒危植物梵净山冷杉野生种群结构及动态特征. 西北植物学报, 2011, 31(7): 1479-1486. |
| [24] | 王卓, 黄荣风, 王林和, 张国盛. 毛乌素沙地天然臭柏种群生命表分析. 中国沙漠, 2009, 29(1): 118-124. |
| [25] | 王电杰, 温国胜, 童在康, 许基全, 邬金, 刘婷霞. 浙江省大鹿岛木麻黄种群静态生命表分析. 内蒙古农业大学学报: 自然科学版, 2011, 32(4): 145-150. |