 2014, Vol. 34
2014, Vol. 34文章信息
- 李玉, 康晓明, 郝彦宾, 丁恺, 王艳芬, 崔骁勇, 梅旭荣
- LI Yu, KANG Xiaoming, HAO Yanbin, DING Kai, WANG Yanfen, CUI Xiaoyong, MEI Xurong
- 黄河三角洲芦苇湿地生态系统碳、水热通量特征
- Carbon,water and heat fluxes of a reed (Phragmites australis) wetland in the Yellow River Delta, China
- 生态学报, 2014, 34(15): 4400-4411
- Acta Ecologica Sinica, 2014, 34(15): 4400-4411
- http://dx.doi.org/10.5846/stxb201212051750
-
文章历史
- 收稿日期:2012-12-5
- 网络出版日期:2014-3-3






2. 中国林业科学研究院湿地研究所, 北京 100091;
3. 中国农业科学院农业环境与可持续发展研究所, 北京 100081
2. Institute of Wetland Research, Chinese Academy of Forestry, Beijing 100091, China;
3. Institute of Environment and Sustainable Development in Agriculture, Beijing 100081, China
全球碳循环中,湿地生态系统作为全球生态系统的重要类型,其碳循环及碳收支的动态变化研究在全球碳收支平衡中扮演着重要角色[1]。湿地作为陆地生态系统最重要的碳库之一,虽然仅占陆地表面的6%左右[2],但其土壤碳储量占陆地土壤总碳储量的10%—30%[3],其碳贮存能够消减大气日益增加的CO2,在稳定全球气候、减缓温室效应方面发挥着重要作用[4]。然而,近年来由于气候变化和人类活动的干扰,湿地面积大幅度萎缩,其正常的生态系统碳循环过程也发生了巨大的变化。因此,研究不同湿地的碳收支状况及其运行机制对精确估算全球碳收支具有重要意义[5, 6]。
目前已有的关于湿地方面的碳源/汇的研究尚无统一的结论。大部分的研究结果证明湿地生态系统是碳汇[7, 8, 9, 10],但随着气候环境变化以及人类活动的影响,部分湿地呈现出碳源的现象[11, 12, 13]。芦苇湿地作为世界上分布最广泛的湿地类型之一,在湿地碳收支的研究中占有重要的地位[14]。已经开展的关于芦苇湿地的研究极少采用直接观测CO2在大气和湿地之间的交换量,这就造成了无法定量分析CO2 交换量与环境因子之间变化的相关关系[14, 15]。而涡度相关技术(Eddy covariance technology)使精确测定湿地生态系统的碳交换成为可能。近年来,涡度相关技术已经成为直接测定大气与群落CO2交换通量的主要方法,也是世界上CO2和水热通量测定的标准方法,所观测的数据已经成为检验各种模型估算精度的最权威的资料。该方法已经得到微气象学家和生态学家的广泛认可,成为目前通量观测网络FlUXNET的主要技术手段,已经在世界范围内被广泛用来测量大气和地球表面碳、水、热通量的交换[16, 17, 18],用这种微气象学方法观测到的净生态系统CO2交换(Net ecosystem CO2 exchange,NEE)能够为在生态系统尺度上了解光合、呼吸提供重要信息[19, 20]。
国外关于湿地碳的研究开展较早且广泛,主要包括碳的累积与分配、碳的分布特征及其影响因素等方面[21]。我国有关湿地碳数量、分布、行为及其动态变化特征的研究主要集中在青藏高原东部的若尔盖高原[22]、东北的三江平原[23]和长江中下游湿地[24, 25, 26, 27],但对黄河三角洲湿地生态系统碳元素生物地球化学循环研究极少。黄河三角洲湿地是典型的滨海湿地,是陆地-海洋-大气相互作用最活跃的地带,是全球环境变化的缓冲区[28]。因此,准确评估黄河三角洲湿地生态系统碳库的源/汇功能及其环境影响机制,将有助于预测全球气候变化与湿地生态系统之间的反馈关系以及湿地资源的可持续利用。然而,由于黄河三角洲滨海湿地生态系统中碳元素的生物地球化学过程十分复杂,各种物理、化学和生物过程共同控制着有机质在湿地中的储存与转化[28, 29, 30],目前对滨海湿地生态系统碳蓄积、碳循环的许多物理、化学和生理生态学过程的理解还十分有限,很多过程的机理尚不完全清楚。而且,湿地生态系统表现出较大的年际间总生产力的波动,这使得在确定湿地生态系统碳库容量及碳源/汇方面依然存在诸多不确定性。因此,在全球气候变化背景下,黄河三角洲芦苇湿地生态系统是碳源还是碳汇?其碳源/汇功能及碳循环动态的控制与反馈机制是什么?这些就成为人们关注的重要科学问题。为了更深入地了解黄河三角洲滨海湿地碳的生物地球化学循环特征及其关键机制,本研究选取黄河三角洲芦苇湿地作为研究对象,结合涡度相关技术,利用长期的通量观测数据和生物量等野外监测数据,探讨生态系统尺度芦苇湿地净生态系统CO2交换量的季节变异特征及其环境控制机制,希望能为区域的碳收支预算和为全球碳循环模型的进一步完善提供理论基础,为重新评价芦苇湿地对全球变化的贡献提供重要的科学依据。
1 实验方法 1.1 研究区概况研究区位于黄河三角洲国家自然保护区(37°40′°N—38°10′N,118°41′E—119°16′E)。该湿地自然保护区地处我国山东省东营市黄河入海口,总面积15.3×104hm2,是以保护黄河口新生湿地生态系统和珍稀濒危鸟类为主体的自然保护区。研究区四季分明,属北温带亚湿润气候区,年平均气温12.1 ℃,年均降水量551.6 mm,无霜期196 d,年蒸发量为1962 mm。土壤类型为潮土、盐土和滨海盐土,土壤表层多以轻沙壤土和沙壤土为主。有机质含量一般在0.6%—1.0% 之间,土壤pH值为7.6—8.5。主要植被有芦苇(Phragmites australis)、穗状狐尾藻 (Myriophyllum spicatum)、荻(Triarrhena sacchariflora)、 蒲草(Typha angustifolia)、补血草(Limonium sinense)、翅碱蓬(Suaeda salsa)、柽柳(Tamarix chinensis)等,芦苇、翅碱蓬和柽柳分布较广[29, 30]。
1.2 观测方法与数据处理通量观测设备主要包括一套开路涡度相关系统和一套常规气象要素测量系统。涡度相关系统主要测量离地面4.5 m高的CO2通量、潜热和感热通量,由一个开路远红外CO2/H2O气体分析仪(IRGA,LI 7500,LI-COR Inc. NE,USA)和一个三维超声波测风仪(CSAT3,Campbell Scientific,MS,USA)组成。仪器采样频率为10 Hz,每半小时自动将平均值记录在数据采集器中(CR5000,Campbell Scientific)。
常规气象要素测量系统包括安装在离地面1.5 m的辐射测定仪(CNR-1,Kipp & Zonen,NY,USA)和光量子测定仪(LI190SB,Li-COR,Lincoln,NE,USA),用于测量净辐射和光合有效辐射。同时在离地面4.5 m处测量相对湿度(HMP45C,Vaisala,Woburn,MA,USA)和风速。土壤温度(地面以下0.05,0.10,0.20,0.5,1.0 m)、土壤热通量(0.05 m,HFP01,HUKESEFLUX,Delft,Netherlands),降水量等要素也同时监测。每半小时输出1组平均值记录在数据采集器中。
为了减少因观测引起的不确定性,我们对数据进行了质量控制和处理。利用涡度相关数据处理软件(Edire软件)对数据进行坐标轴旋转和WPL校正,以消除地形倾斜对通量计算的影响,同时也校正了由于空气水热传输引起的CO2和水汽密度波动造成的通量计算误差[31, 32, 33]。由于降雨、标定和仪器故障(如系统维护、电压不稳、断电等)等原因必然会造成数据缺失和一些异常点的出现。同时,较低的摩擦风速(u*)和夜间低湍流也会低估系统的净CO2交换速率[34, 35]。为了消除这些影响,对数据进行了筛选和质量控制,大约30%的数据被剔除掉。具体处理过程见Hao等[36]。白天缺失的NEE数据通过Michaelis-Menten方程法[37]插补得到,
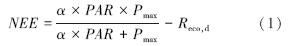
式中,α(mg CO2 μmol 光子-1)为生态系统表观光量子效率,Pmax为潜在最大CO2同化速率,Reco,d(mg CO2 m-2 s-1)是白天的生态系统呼吸速率,PAR(μmol m-2 s-1)是光合有效辐射。
由于夜间湍流不充分,剔除u*(摩擦风速)<0.15 m/s时的NEE数据,对夜间缺失数据通过呼吸模型[39]得到:

式中,a,b是常数,Ts是5 cm土壤温度,Reco,n (mg CO2 m-2 s-1)是夜间的生态系统呼吸速率。
对插补后的数据划分白天(总辐射≥l W·m )和晚上(总辐射<l W·m )。选取每个月晴朗无云天气下的数据作代表,分析整个生长季NEE和其它变量的日动态变化;并且把白天和晚上每30 min的通量进行积分求和,分别代表生态系统白天和晚上的CO2累积吸收量和累积释放量。
通量塔直接观测到的CO2通量代表了净生态系统CO2交换(NEE),正值代表系统净排放CO2,负值代表系统净吸收CO2,是生态系统呼吸(Re)和总初级生产力(GPP)之间的差值[38]。因此,GPP可以定义为:

日生态系统呼吸是白天生态系统呼吸(Re,day)和夜间生态系统呼吸的总和(Re,night):
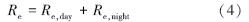
夜间生态系统呼吸由观测到的夜间净CO2 通量得到。用呼吸模型(方程2)外推白天的生态系统呼吸[39]。
1.3 地上生物量和叶面积测量地上生物量的测定采用刈割收获法。将样方内的芦苇齐地面剪下,带回实验室,称其鲜重后65℃烘干至恒重。从2010年6月到9月每月测定1次,每次取4个1 m×1 m的样方。
叶面积指数采用称重法,先在硫酸纸上画出10 cm×10 cm的正方形,用剪刀剪下称重,求出平均1 cm2的纸重WP(g),作为计算叶面积的常数,然后将叶片平铺在同一种纸上,用硬铅笔绘出叶片的轮廓,剪刀剪下纸的轮廓,称重WL(g),叶面积A(cm2)然后根据公式即可求出LAI。
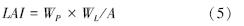
本研究利用路径分析的方法来评价2009—2010年各个环境因子对CO2通量的影响。路径分析已经被广泛用于评价多个环境变量对碳通量季节和年际动态的相对重要性[40, 41, 42]。这种方法是一种增强型的多元回归分析,能够用来评价各个环境变量之间的相互关系及对碳通量的直接和间接影响程度。本研究主要关注4个环境因子(空气温度(Ta)、土壤温度(Ts)、降水量(PPT)和光合有效辐射(PAR))对芦苇湿地CO2通量的影响。对于建立的模型,利用3个拟合度指数进行模型拟合度评估,若模型拟合度越高,则代表模型可用性越高,参数的估计越具有其涵义。3个拟合度指数分别为卡方统计量(x2)、基准化适合度指标(NFI,normed fit index)和比较适合度指标(CFI,comparative fit index)。其中,x2一般以卡方值P > 0.05作为判断,意即模型具有良好的拟合度;NFI和CFI越接近于1表示模型拟合度越好。路径分析的软件是AMOS 20.0 (Analysis of Moment Structures)。
2 结果 2.1 环境和生物因子研究期间,环境和生物因子都表现出明显的季节变化。2009年年均温为13.9 ℃,略高于2010年(13.3 ℃),但都高于多年平均气温(12.1 ℃)(图 1)。两年的最高气温均为33.4 ℃,分别出现在6月25日和7月30日;最低气温均出现在1月份。2009—2010年的降水表现出很大的季节波动和年际波动(图 1)。2009年降水量为571.4 mm,高于多年平均降水量(551 mm)。而2010年降水量为523.5 mm,低于多年平均降水量。2009年各月降水除1、8、9月3个月,其余月份均比2010年高。2009年降水主要集中在5、6、7月3个月,占全年总降水量的71.9%。2010年降水主要集中在8月份,单月降水301.7 mm,占全年总降水量的58%。

|
| 图 1 黄河三角洲芦苇湿地2009—2010年空气温度和降水量的季节变化特征 Fig. 1 Seasonal variations of air temperature (Ta) and precipitation (PPT) during 2009—2010 in the Yellow River Delta |
地上生物量和叶面积指数均在8月达到最大值,分别为(433.33±57.16)g/m2和(2.22±0.67)m2/m2。7月份地上生物量比6月份有所减少,8月份急剧增加,很可能是由于7月降水较少(33.1 mm),而从8月开始降水增加,有几次强降水,月总降水量为301.7 mm,占2010年总降水量的58%;到9月,降水再次减少,仅为26.5 mm,故9月份地上生物量急剧下降,植物开始逐渐枯萎(图 2)。

|
| 图 2 2010年地上生物量和叶面积指数的季节动态 Fig. 2 Seasonal variations of the above-ground biomass and the leaf area index (LAI) in 2010 |
光合有效辐射(PAR)是控制光合作用的主要因素之一,因此PAR的大小也强烈影响NEE的大小。图 3给出了2009—2010年生长季芦苇湿地净生态系统碳交换(NEE)的日动态。生态系统CO2通量的日进程表现出两种不同的类型。一种类型是生态系统碳通量具有两个吸收高峰。以晴朗天气半小时碳通量平均值为例,日出后(大约7:00左右)生态系统开始吸收CO2,随着PAR的增加,光合作用逐渐增强,湿地NEE逐渐由净排放(正值)转为净吸收(负值),固碳速率逐渐增大,大约在10:00—11:00左右达到CO2吸收峰值。但随后随着PAR的继续增加,系统固碳速率开始逐渐降低(图 3,图 4)。14:00左右生态系统固碳能力降低,出现“午休”现象。到午后16:00左右,生态系统固碳能力又开始增强,达到第2个CO2吸收高峰,19:00左右NEE接近于0,生态系统开始向大气中排放CO2。另一种类型是生态系统只有1个吸收高峰(图 3)。日出后,生态系统开始吸收CO2,固碳速率随着PAR的增加逐渐增大,12:00—13:00左右达到吸收高峰后,生态系统的固碳速率开始逐渐下降,19:00—20:00左右,生态系统转变为CO2的源。通过对芦苇湿地生态系统连续2a的观测数据分析发现,CO2的最大吸收速率为0.30 和0.37 mg CO2 m-2 s-1,分别出现在2009年7月的11:00左右和2010年6月的13:00左右。CO2释放的最大值均出现在9月,分别为0.19和0.25 mg CO2 m-2s-1(图 3)。

|
| 图 3 2009—2010年净生态系统碳交换的日动态 Fig. 3 The diurnal patterns of the net ecosystem CO2 exchange (NEE) in 2009 and 2010 |

|
| 图 4 2009年和2010年生长季各月光合有效辐射和净生态系统碳交换的关系 Fig. 4 The relationship between photosynthetic active radiation (PAR) and the net ecosystem CO2 exchange (NEE) in each month of growing season in 2009 and 2010 |
2009、2010年生长季各月感热通量的平均日动态曲线呈单峰型(图 5),白天由于太阳辐射的原因 导致湿地表面温度高于大气温度,感热通量向上运输,在12:00左右达到最大值;夜晚由于湿地地表辐射减少形成近地面逆温层,湿地表面温度低于大气温度,感热通量向下运输。从各月的感热通量日动态来看,两年的7、8、9三个月之间的变化幅度都不大。潜热通量的平均日动态曲线也呈单峰型(图 5)。

|
| 图 5 2009—2010年潜热通量和感热通量的日动态 Fig. 5 The diurnal patterns of the sensible heat fluxes (Hs) and the latent heat fluxes (LE) in 2009 and 2010 |
两年生长季感热通量比较结果表明,2010 年除7 月份白天略高于2009 年同期外,其余各月份均小于2009 年同期,而潜热通量的比较表明,2010 年8 月份潜热通量低于2009年同期,其余各月高于2009 年同期。由波文比日动态(图 6)可以看出,大部分|Hs/LE| < 1,潜热占主导地位,只有2009 年10 月和2010 年9 月夜间感热大于潜热。白天,波文比为正值,最大值分别为0.82(2009 年10 月)和0.52(2010 年7 月)。夜晚感热、潜热数值较小,且易受影响,故波动比较大。2010 年9 月由于降水骤减,导致当月感热通量增加,波文比波动剧烈。

|
| 图 6 2009—2010年波文比日动态 Fig. 6 The diurnal pattern of Bowen ratio in 2009 and 2010 |
从热通量和CO2通量的相关性分析(图 7)得出,潜热通量、感热通量均和NEE有显著的负相关关系,即一天中随着热通量的增加,NEE值减小,且感热通量和NEE的相关性高于潜热通量和NEE的相关性,R2值分别为0.50和0.59。

|
| 图 7 热通量和NEE的关系 Fig. 7 The relationship between heat fluxes and NEE |
2009年芦苇湿地CO2通量累积白天最大吸收值出现在9月2日,为-9.38 g CO2 m-2 d-1,夜晚最大排放值出现在9月3日,为6.49 g CO2 m-2 d-1(图 8)。从季节动态分析,芦苇湿地在7、8月份生态系统CO2净吸收能力最强,从9月份开始,吸收能力开始逐渐下降。2009年生长季生态系统白天固定-354.63 g CO2/m2,同时期夜间释放了159.24 g CO2/m2,净CO2交换为-195.39g CO2/m2 (负值代表碳吸收)。2009年生长季生态系统总初级生产力 (GPP)最大值出现在9月2日,为-11.90 g CO2 m-2 d-1,最小值出现在9月22日,为1.56 g CO2 m-2 d-1;生态系统呼吸(Re)最大值出现在9月3日,为9.05 g CO2 m-2 d-1,最小值出现在10月10日,为2.06 g CO2 m-2 d-1(图 9)。2009年整个生长季生态系统GPP为-651.13 g CO2/m2,Re为455.74 g CO2/m2,系统表现为碳汇。

|
| 图 8 2009年生长季净生态系统碳交换 (NEE) 白天累积值 (NEEday) ,夜晚累积值 (NEEnight) 和日总累计值 (NEEtotal) 季节动态 Fig. 8 The seasonal dynamics of integrated daytime,nighttime and total NEE in 2009 growing season |

|
| 图 9 2009年生长季碳通量日累积季节变化 (总初级生产力GPP,净生态系统碳交换NEE,生态系统呼吸Re) Fig. 9 The seasonal dynamics of integrated carbon fluxes (GPP,NEE,Re) in in 2009 growing season |
在芦苇湿地生态系统,多个环境因子会影响NEE通量。本研究利用路径分析的方法探讨各个环境因子对芦苇湿地碳收支的直接影响和间接影响(图 10)。模型的拟合度非常好,NFI和CFI都>0.9,模型可用性极高。在直接影响NEE通量的3个环境因子中(Ts,PAR和PPT),Ts对NEE通量的路径系数是-0.52,明显高于PPT和PAR与NEE通量的之间的路径系数(分别是0.26和-0.12)(图 10)。2a的观测结果表明,土壤温度是黄河三角洲芦苇湿地CO2通量变化的主要影响因子,且土壤温度、光合有效辐射与NEE呈显著的负相关关系,降水量与NEE呈显著的正相关关系,即随着PAR的增强、土壤温度的升高及降水量的减少,芦苇湿地生态系统的固碳能力在逐渐增强。而且,路径分析结果发现,Ta对Ts有显著的影响,这表明空气温度通过影响土壤温度间接影响NEE通量。Ta对NEE的间接影响为-0.43,表明空气温度与NEE通量为负相关关系。其它环境因子对NEE的间接影响非常小。

|
| 图 10 2009—2010年路径分析图,表明各个环境因子 (空气温度Ta,0—5 cm土壤温度Ts,光合有效辐射PAR和降水量PPT) 对NEE通量的直接和间接影响 Fig. 10 Path diagrams illustrating the effects of environmental variables (air temperature (Ta),soil temperature at a depth of 5 cm (Ts),photosynthetically active radiation (PAR) and precipitation (PPT)) on net ecosystem CO2 exchange (NEE) during 2009—2010 每个箭头的粗细和箭头上的数值代表路径系数及其大小,实线代表负相关,虚线代表正相关 |
通过对黄河三角洲芦苇湿地2009、2010两年生长季通量数据分析,得到以下结论:
(1) 芦苇湿地具有较强的碳汇功能。2009年生长季共吸收-195.39 g CO2 m-2,低于同年若尔盖湿地的固碳能力(-292.23 g CO2 m-2)[22],也低于温带草甸湿地[10](-199 g C m-2 a-1)和温带香蒲沼泽[9](强碳汇-264 g C m-2 a-1)的固碳潜势,但要高于青藏高原东北部的乱海子湿地[12](弱碳源24.1 g C m-2)。CO2通量日变化表现为白天吸收,夜晚排放。黄河三角洲芦苇湿地和国外及国内芦苇湿地比较,其生态系统碳通量的大体特征一致,都为碳汇[6, 43, 44, 45, 46],但其日动态值低于若尔盖高寒湿地(-0.54 mg CO2 m-2 s-1)和辽河三角洲芦苇湿地(-1.24 mg CO2 m-2 s-1),故三角洲芦苇湿地碳汇功能弱于其它同类型的湿地系统。
(2) 芦苇湿地夏季由于高温、强光、低湿等环境条件引起部分气孔关闭,或光合作用被抑制[47, 48],在14:00左右出现“午休”现象,时间晚于草原生态系统(例如内蒙古羊草草原8:00—10:00,青海湖草甸草原11:30—13:00)[13, 49, 50]。植物光合作用过程中这种午间降低现象已被许多研究证实,是较为普遍的现象[47, 48, 50, 51]。而黄河三角洲芦苇湿地,由于其外在气候、地理环境条件及自身生理条件与草原生态系统的差异[13, 30, 52],导致其“午休”现象出现时间的推迟。
(3) 芦苇湿地的潜热通量始终为正,反映了芦苇湿地较强的水循环特性,芦苇湿地全天水汽均向上输送[43]。感热潜热的季节变化受温度和降水的综合影响。生长季只有部分月份表现为白天潜热通量占主导,夜晚感热通量占主导,和周广胜等对盘锦芦苇湿地能量通量的研究结论一致[43],而这一研究结果很好地验证了降水对波文比影响显著,干旱会导致感热通量增加,甚至超过潜热通量[53]。而在日尺度上,潜热通量、显热通量和NEE有显著的负相关关系,这也验证了水热和CO2耦合模型中关于水热传输和CO2传输过程的模拟机理[54]。
(4) 光合有效辐射(PAR)和温度是影响NEE通量的主要环境因子。在日尺度上,NEE和PAR值之间表现出明显的双曲线关系,这种关系已被广泛证实[6, 52, 55, 56]。从季节动态来看,由路径分析发现土壤温度是黄河三角洲芦苇湿地CO2通量变化的主要影响因子,而降水量和PAR对CO2通量的变化影响次之,这和Hao 等对若尔盖高原高寒湿地的研究结论基本一致,认为PAR、温度和降水显著影响湿地生态系统的CO2通量[22]。其它湿地的研究也发现了温度在控制碳平衡中的重要性[57, 58, 59]。而在高海拔或者高寒地区,昼夜温差以及温度的季节变化非常大,因此也成为控制CO2通量的重要环境变量[22, 60]。然而,这些研究缺乏各个环境因子对CO2通量相对重要性和贡献量的比较研究。本研究的路径分析则提供了一些新的信息,明确反映了温度及其它环境因子对若尔盖湿地碳通量的直接影响、间接影响及影响程度。
| [1] | Huntingford C, Lowe J A, Booth B B B, Jones C D, Harris G R, Gohar L K, Meir P. Contributions of carbon cycle uncertainty to future climate projection spread. Tellus B, 2009, 61(2): 355-360. |
| [2] | Gao J Q, Xu X L, Zhang F, Wang C M. Distribution characteristics of soil labile carbon along water table gradient of alpine wetland soils. Journal of Soil and Water Conservation, 2008, 22(3): 126-131. |
| [3] | Smith L C, MacDonald G M, Velichko A A, Beilman D W, Borisova O K, Frey K E, Kremenetski K V, Sheng Y. Siberian peatlands a net carbon sink and global methane source since the early Holocene. Science, 2004, 303(5656): 353-356. |
| [4] | Yu G R. Global Change and Terrestrial Ecosystem Carbon Cycle and Carbon Accumulation. Beijing: Meteorological Press, 2003: 43-96. |
| [5] | Song C C. Advance in research on carbon cycling in wetlands. Scientia Geographica Sinica, 2003, 23(5): 622-628. |
| [6] | Zhou L, Zhou G S, Jia Q Y. Annual cycle of CO2 exchange over a reed (Phragmites australis) wetland in Northeast China. Aquatic Botany, 2009, 91(2): 91-98. |
| [7] | Suyker A E, Verma S B, Arkebauer T J. Season-long measurement of carbon dioxide exchange in a boreal fen. Journal of Geophysical Research, 1997, 102(D24): 29021-29028. |
| [8] | Lafleur P M, Roulet N T, Bubier J L, Frolking S, Moore T R. Interannual variability in the peatland-atmosphere carbon dioxide exchange at an ombrotrophic bog. Global Biogeochemical Cycles, 2003, 17(2): 1036-1049. |
| [9] | Bonneville M C, Strachan I B, Humphreys E R, Roulet N T. Net ecosystem CO2 exchange in a temperate cattail marsh in relation to biophysical properties. Agricultural and Forest Meteorology, 2008, 148(1): 69-81. |
| [10] | Dušek J, Č ížková H, Czerný R, Taufarová K, Šmídová M, Janouš D. Influence of summer flood on the net ecosystem exchange of CO2 in a temperate sedge-grass marsh. Agricultural and Forest Meteorology, 2009, 149(9): 1524-1530. |
| [11] | Griffis TJ, Rouse W R, Waddington J M. Scaling net ecosystem CO2 exchange from the community to landscape-level at a subarctic fen. Global Change Biology, 2000, 6(4): 459-473. |
| [12] | Hirota M, Tang Y H, Hu Q W, Hirata S, Kato T, Mo W H, Cao G M, Mariko S. Carbon dioxide dynamics and controls in a deep-water wetland on the Qinghai-Tibetan Plateau. Ecosystems, 2006, 9(4): 673-688. |
| [13] | Zhang F W, Liu A H, Li Y N, Zhao L, Wang Q X, Du M Y. CO2 flux in alpine wetland ecosystem on the Qinghai-Tibetan Plateau, China. Acta Ecologica Sinica, 2008, 28(2): 453-462. |
| [14] | Brix H, Sorrell B K, Lorenzen B. Are Phragmites-dominated wetlands a net source or net sink of greenhouse gases?. Aquatic Botany, 2001, 69(2/4): 313-324. |
| [15] | Huang G H, Li X Z, Hu Y M, Shi Y, Xiao D N. Methane (CH4) emission from a natural wetland of Northern China. Journal of Environmental Science and Health, 2005, 40(6/7): 1227-1238. |
| [16] | Baldocchi D, Falge E, Gu L H, Olson R, Hollinger D, Running S, Anthoni P, Bernhofer C, Davis K, Evans R, Fuentes J, Goldstein A, Katul G, Law B, Lee X H, Malhi Y, Meyers T, Munger W, Oechel W, Paw K T, Pilegaard K, Schmid H P, Valentini R, Verma S, Vesala T, Wilson K, Wofsy S. FLUXNET: A new tool to study the temporal and spatial variability of ecosystem-scale carbon dioxide, water vapor, and energy flux densities. Bulletin of the American Meteorological Society, 2001, 82(11): 2415-2434. |
| [17] | Hutley L B, Leuning R, Beringer J, Cleugh H A. The utility of the eddy covariance techniques as a tool in carbon accounting: tropical savanna as a case study. Australian Journal of Botany, 2005, 53(7): 663-675. |
| [18] | Kang X M, Hao Y B, Li C S, Cui X Y, Wang J Z, Rui Y C, Niu H S, Wang Y F. Modeling impacts of climate change on carbon dynamics in a steppe ecosystem in Inner Mongolia, China. Journal of Soils and Sediments, 2011, 11(4): 562-576. |
| [19] | Falge E, Baldocchi D, Tenhunen J, Aubinet M, Bakwin P, Berbigier P, Bernhofer C, Burba G, Clement R, Davis K J, Elbers J A, Goldstein A H, Grelle A, Granier A, Guömundsson J, Hollinger D, Kowalski A S, Katul G, Law B E, Malhi Y, Meyers T, Monson R K, Munger J W, Oechel W, Paw K T, Pilegaard K, Rannik Ü, Rebmann C, Suyker A, Valentini R, Wilson K, Wofsy S. Seasonality of ecosystem respiration and gross primary production as derived from FLUXNET measurements. Agricultural and Forest Meteorology, 2002, 113(1/4): 53-74. |
| [20] | Sanderman J, Amundson R G, Baldocchi D D. Application of eddy covariance measurements to the temperature dependence of soil organic matter mean residence time. Global Biogeochemical Cycles, 2003, 17(2), doi: 10.1029/2001GB001833. |
| [21] | Rhee J S, Iamchaturapatr J. Carbon capture and sequestration by a treatment wetland. Ecological Engineering, 2009, 35(3): 393-401. |
| [22] | Hao Y B, Cui X Y, Wang Y F, Mei X R, Kang X M, Wu N, Luo P, Zhu D. Predominance of precipitation and temperature controls on ecosystem CO2 exchange in zoige alpine wetlands of Southwest China. Wetlands, 2011, 31(2): 413-422. |
| [23] | Xi M, Lu X G, Li Y, Kong F L. Distribution characteristics of dissolved organic carbon in annular wetland soil-water solutions through soil profiles in the Sanjiang Plain, Northeast China. Journal of Environmental Sciences, 2007, 19(9): 1074-1078. |
| [24] | Zhou J L, Wu Y, Kang Q S, Zhang J. Spatial variations of carbon, nitrogen, phosphorous and sulfur in the salt marsh sediments of the Yangtze Estuary in China. Estuarine, Coastal and Shelf Science, 2007, 71(1/2): 47-59. |
| [25] | Yan Y, Zhao B, Chen J Q, Guo H Q, Gu Y J, Wu Q H, Li B. Closing the carbon budget of estuarine wetlands with tower-based measurements and MODIS time series. Global Change Biology, 2008, 14(7): 1690-1702. |
| [26] | Yan Y E, Guo H Q, Gao Y, Zhao B, Chen J Q, Li B, Chen J K. Variations of net ecosystem CO2 exchange in a tidal inundated wetland: Coupling MODIS and tower-based fluxes. Journal of Geophysical Research, 2010, 115: D15102. |
| [27] | Guo H Q, Noormets A, Zhao B, Chen J Q, Sun G, Gu Y J, Li B, Chen J K. Tidal effects on net ecosystem exchange of carbon in an estuarine wetland. Agricultural and Forest Meteorology, 2009, 149(11): 1820-1828. |
| [28] | Yu J B, Chen X B, Sun Z G, Xie W J, Mao P L, Wu C F, Dong H F, Mu X J, Li Y Z, Guan B, Shan K. The spatial distribution characteristics of soil nutrients in new-born coastal wetland in the Yellow River delta. Acta Scientiae Circumstantiae, 2010, 30(4): 855-861. |
| [29] | Dong H F, Yu J B, Sun Z G, Mu X J, Chen X B, Mao P L, Wu C F, Guan B. Spatial distribution characteristics of organic carbon in the soil-plant systems in the Yellow River estuary tidal flat wetland. Chinese Journal of Environmental Science, 2010, 31(6): 1594-1599. |
| [30] | He Q, Cui B S, Zhao Y S, Fu H L, Xiong X, Feng G H. Vegetation distribution patterns to the gradients of water depth and soil salinity in wetlands of Yellow River Delta, China. Wetland Science, 2007, 5(3): 208-214. |
| [31] | Wilczak J M, Oncley S P, Stage S A. Sonic anemometer tilt correction algorithms. Boundary-Layer Meteorology, 2001, 99(1): 127-150. |
| [32] | Zhu Z L, Sun X M, Wen X F, Zhou Y L, Tian J, Yuan G F. ChinaFLUXE night CO2 eddy covariance flux data processing method research. Science in China (D): Earth Science, 2006, 36(Supplement 1): 34-44. |
| [33] | Webb E K, Pearman G I, Leuning R. Correction of flux measurements for density effects due to heat and water vapour transfer. Quarterly Journal of the Royal Meteorological Society, 1980, 106(447): 85-100. |
| [34] | Goulden M L, Munger J W, Fan S M, Daube B C, Wofsy S C. Measurements of carbon sequestration by long-term eddy covariance: methods and a critical evaluation of accuracy. Global Change Biology, 1996, 2(3): 169-182. |
| [35] | Jaksic V, Kiely G, Albertson J, Oren R, Katul G, Leahy P, Byrne K A. Net ecosystem exchange of grassland in contrasting wet and dry years. Agricultural and Forest Meteorology, 2006, 139(3/4): 323-334. |
| [36] | Hao Y B, Wang Y F, Huang X Z, Cui X Y, Zhou X Q, Wang S P, Niu H S, Jiang G M. Seasonal and interannual variation in water vapor and energy exchange over a typical steppe in Inner Mongolia, China. Agricultural and Forest Meteorology, 2007, 146(1/2): 57-69. |
| [37] | Falge E, Baldocchi D, Olson R, Anthoni P, Aubinet M, Bernhofer C, Burba G, Ceulemans R, Clement R, Dolman H, Granier A, Gross P, Grünwald T, Hollinger D, Jensen N O, Katul G, Keronen P, Kowalski A, Lai C T, Law B E, Meyers T, Moncrieff J, Moors E, Munger J W, Pilegaard K, Rannik V, Rebmann C, Suyker A, Tenhunen J, Tu K, Verma S, Vesala T, Wilson K, Wofsy S. Gap filling strategies for defensible annual sums of net ecosystem exchange. Agricultural and Forest Meteorology, 2001, 107(1): 43-69. |
| [38] | Law B E, Falge E, Gu L, Baldocchi D D, Bakwin P, Berbigier P, Davis K, Dolman A J, Falk M, Fuentes J D, Goldstein A, Granier A, Grelle A, Hollinger D, Janssens I A, Jarvis P, Jensen N O, Katul G, Mahli Y, Matteucci G, Meyers T, Monson R, Munger W, Oechel W, Olson R, Pilegaard K, Paw K T, Thorgeirsson H, Valentini R, Verma S, Vesala T, Wilson K, Wofsy S. Environmental controls over carbon dioxide and water vapor exchange of terrestrial vegetation. Agricultural and Forest Meteorology, 2002, 113(1/4): 97-120. |
| [39] | Lioyd J, Taylor J A. On the temperature dependence of soil respiration. Functional Ecology, 1994, 8(3): 315-323. |
| [40] | Wang Y F, Cui X Y, Hao Y B, Mei X R, Yu G R, Huang X Z, Kang X M, Zhou X Q. The fluxes of CO2 from grazed and fenced temperate steppe during two drought years on the Inner Mongolia Plateau, China. Science of the Total Environment, 2011, 410-411: 182-190. |
| [41] | Saito M, Kato T, Tang Y H. Temperature controls ecosystem CO2 exchange of an alpine meadow on the northeastern Tibetan Plateau. Global Change Biology, 2009, 15(1): 221-228. |
| [42] | Matías L, Castro J, Zamora R. Effect of simulated climate change on soil respiration in a mediterranean-type ecosystem: rainfall and habitat type are more important than temperature or the soil carbon pool. Ecosystems, 2012, 15(2): 299-310. |
| [43] | Wang H Y, Zhou G S. Long-term flux characteristics of Phragmites ecosystem in Panjin wetland. Journal of Meteorology and Environment, 2006, 22(4): 18-24. |
| [44] | Suzuki S, Yokozawa M, Inubushi K, Hara T, Kimura M, Tsuga S, Tako Y, Nakamura Y. Evaluation of CO2 exchange rates in a wetland ecosystem using the closed geosphere experiment facility. Journal of Hydrometeorology, 2012, 13(3): 966-980. |
| [45] | Mander Ü, Lõhmus K, Teiter S, Mauring T, Nurk K, Augustin J. Gaseous fluxes in the nitrogen and carbon budgets of subsurface flow constructed wetlands. Science of the Total Environment, 2008, 404(2/3): 343-353. |
| [46] | Milla R, Cornelissen J H C, van Logtestijn R S P, Toet S, Aerts R. Vascular plant responses to elevated CO2 in a temperate lowland Sphagnum peatland. Plant Ecology, 2006, 182(1/2): 13-24. |
| [47] | Xu D Q. Photosynthetic "nap" phenomenon. Plant Physiology Journal, 1997, (6): 466-467. |
| [48] | Xu D Q. Photoinhibition of photosynthesis. Plant Physiology Journal, 1997, (6): 467-467. |
| [49] | Hao Y B, Wang Y F, Sun X M, Huang X Z, Cui X Y, Niu H S, Zhang Y H, Yu G R. Seasonal variation in carbon exchange and its ecological analysis over Leymus chinensis steppe in Inner Mongolia. Science in China (D): Earth Science, 2006, 36(Supplement 1): 174-182. |
| [50] | Shi S B, Han F, Li H Y. Midday depression of photosynthesis of Gentiana straminea and Saussurea superba in Alpine Kobresia humilis Meadow. Acta Phytophysiologica Sinica, 2001, 27(2): 123-128. |
| [51] | Guehl J M, Aussenac G. Photosynthesis decrease and stomatal control of gas exchange in Abies alba mill. in response to vapor pressure difference. Plant Physiology, 1987, 83(2): 316-322. |
| [52] | Hao Y B, Wang Y F, Mei X R, Huang X Z, Cui X Y, Zhou X Q, Niu H S. CO2, H2O and energy exchange of an Inner Mongolia steppe ecosystem during a dry and wet year. Acta Oecologica, 2008, 33(2): 133-143. |
| [53] | Yin D Z, Chen J Y. Modeling study on the land surface fluxes and the structure of planetary boundary layer (PBL) over Inner Mongolia Grassland. Acta Agrestia Sinica, 1998, 6(4): 282-292. |
| [54] | Wang J, Yu Q, Pan X B, Yin H, Zhang Y Q. A review on water, heat and CO2 fluxes simulation models. Acta Ecologica Sinica, 2008, 28(6): 2843-2853. |
| [55] | Sims P L, Bradford J A. Carbon dioxide fluxes in a southern plains prairie. Agricultural and Forest Meteorology, 2001, 109(2): 117-134. |
| [56] | Huang X Z, Hao Y B, Wang Y F, Zhou X Q, Han X, He J J. Impact of extreme drought on net ecosystem exchange from Lemus chinensis steppe in Xilin River Basin, China. Journal of Plant Ecology, 2006, 30(6): 894-900. |
| [57] | Inglett K S, Inglett P W, Reddy K R, Osborne T Z. Temperature sensitivity of greenhouse gas production in wetland soils of different vegetation. Biogeochemistry, 2012, 108(1/3): 77-90. |
| [58] | Schedlbauer J L, Munyon J W, Oberbauer S F, Gaiser E E, Starr G. Controls on ecosystem carbon dioxide exchange in short- and long-hydroperiod florida everglades freshwater marshes. Wetlands, 2012, 32(5): 801-812. |
| [59] | Xie X, Zhang M Q, Zhao B, Guo H Q. Temperature dependence of coastal wetland ecosystem respiration confounded by tidal activities: a temporal perspective. Biogeosciences Discussions, 2013, 10(3): 4515-4537. |
| [60] | Huxman T E, Turnipseed A A, Sparks J P, Harley P C, Monson R K. Temperature as a control over ecosystem CO2 fluxes in a high-elevation, subalpine forest. Oecologia, 2003, 134(4): 537-546. |
| [2] | 高俊琴, 徐兴良, 张锋, 王春梅. 水分梯度对若尔盖高寒湿地土壤活性有机碳分布的影响. 水土保持学报, 2008, 22(3): 126-131. |
| [4] | 于贵瑞. 全球变化与陆地生态系统碳循环和碳蓄积. 北京: 气象出版社, 2003: 43-96. |
| [5] | 宋长春. 湿地生态系统碳循环研究进展. 地理科学, 2003, 23(5): 622-628. |
| [28] | 于君宝, 陈小兵, 孙志高, 谢文军, 毛培利, 吴春发, 董洪芳, 牟晓杰, 栗云召, 管博, 单凯. 黄河三角洲新生滨海湿地土壤营养元素空间分布特征. 环境科学学报, 2010, 30(4): 855-861. |
| [29] | 董洪芳, 于君宝, 孙志高, 牟晓杰, 陈小兵, 毛培利, 吴春发, 管博. 黄河口滨岸潮滩湿地植物-土壤系统有机碳空间分布特征. 环境科学, 2010, 31(6): 1594-1599. |
| [32] | 朱治林, 孙晓敏, 温学发, 周艳莲, 田静, 袁国富. 中国通量网(ChinaFLUX)夜间CO2涡度相关通量数据处理方法研究. 中国科学D辑: 地球科学, 2006, 36(增刊1): 34-44. |
| [43] | 汪宏宇, 周广胜. 盘锦湿地芦苇生态系统长期通量观测研究. 气象与环境学报, 2006, 22(4): 18-24. |
| [47] | 许大全. 光合作用的“午睡”现象. 植物生理学通讯, 1997, (6): 466-467. |
| [48] | 许大全. 光合作用的光抑制. 植物生理学通讯, 1997, (6): 467-467. |
| [49] | 郝彦宾, 王艳芬, 孙晓敏, 黄祥忠, 崔骁勇, 牛海山, 张亚红, 于贵瑞. 内蒙古羊草草原碳交换季节变异及其生态学解析. 中国科学D辑: 地球科学, 2006, 36(增刊1): 174-182. |
| [50] | 师生波, 韩发, 李红彦. 高寒草甸麻花艽和美丽风毛菊的光合速率午间降低现象. 植物生理学报, 2001, 27(2): 123-128. |
| [53] | 殷达中, 陈家宜. 内蒙古草原陆面通量和边界层结构数值模拟. 草地学报, 1998, 6(4): 282-292. |
| [54] | 王靖, 于强, 潘学标, 尹红, 张永强. 土壤-植物-大气连续体水热、CO2通量估算模型研究进展. 生态学报, 2008, 28(6): 2843-2853. |
| [56] | 黄祥忠, 郝彦宾, 王艳芬, 周小奇, 韩喜, 贺俊杰. 极端干旱条件下锡林河流域羊草草原净生态系统碳交换特征. 植物生态学报, 2006, 30(6): 894-900. |