 2014, Vol. 34
2014, Vol. 34文章信息
- 吴伊波, 车荣晓, 马双, 邓永翠, 朱敏健, 崔骁勇
- WU Yibo, CHE Rongxiao, MA Shuang, DENG Yongcui, ZHU Minjian, CUI Xiaoyong
- 高寒草甸植被细根生产和周转的比较研究
- Estimation of root production and turnover in an alpine meadow:comparison of three measurement methods
- 生态学报, 2014, 34(13): 3529-3537
- Acta Ecologica Sinica, 2014, 34(13): 3529-3537
- http://dx.doi.org/10.5846/stxb201307031831
-
文章历史
- 收稿日期:2013-7-3
- 修订日期:2014-4-18






2. 中国科学院大学生命科学学院, 北京 100049;
3. 南京师范大学虚拟地理环境教育部重点实验室, 南京 210046
2. College of Life Sciences, University of Chinese Academy of Sciences, Beijing 100049, China;
3. Key Laboratory of Virtual Geographic Environment, Ministry of Education, Nanjing Normal University, Nanjing 210046, China
植物根系是陆地生态系统碳及其它矿质元素的重要储存库,细根周转是植被碳库与土壤碳库交换的重要途径之一。大量研究表明,细根生产消耗的碳量占森林生态系统净初级生产力的10%—60%[1, 2];而通过细根周转进入土壤的碳占土壤总输入碳量的14%—50%[3],在有些生态系统甚至是地上凋落物向土壤输入碳量的1.6倍[3- 4]。因此准确估算细根生产及其周转速率是认识生态系统碳分配和养分循环的关键[5, 6, 7],更是正确理解生态系统的碳循环过程及其对全球变化响应的基础[8, 9, 10]。只有准确测定细根周转,才能准确估计通过细根周转和分解向土壤系统输送的碳和养分的总量。
根系研究发展至今,出了多种研究测定方法,但它们都存在着各种不足之处[11, 12],致使研究结果可能出现较大误差[13, 14],对细根生产周转达到准确估计仍非常困难,从而制约了根系研究的进展。由于根系生存于土壤环境中,难以对它们的生理生态活动进行直接观测,加大了研究难度。迄今为止,对根系的研究仍然缺乏统一可靠的采样及分析方法[15, 16, 17, 18, 19, 20, 21],不同研究结果之间缺乏可比性[7]。现有的根系研究报道大多采用传统的方法[22, 23, 24],但近年来微根管实时监测手段逐渐得到了广泛的应用[25, 26, 27]。不同测定方法基于不同的观测原理、假设条件,测定结果存在不同程度的不确定性[11, 28, 29]。而细根周转速率系单位时间内根系生长或死亡的量占根系总生物量的比例,因此其含义与测定方法密切相关。根钻法、生长袋法、微根管法是目前细根研究中应用最广泛的3种方法。根钻法和内生长袋法是通过测定细根生物量随时间的变化来估算细根的生产力,用细根生产力除以其平均(或最大)现存生物量得到细根的周转速率[29];微根管法大都通过记录个体根的存活时间,获得根群的寿命分布,通过统计方法得到中值寿命,而中值寿命的倒数即为细根周转速率[29]。本研究在同一研究地点,采用根钻法、内生长袋法和微根管法开展同一植被类型群落的地下净初级生产力及周转率的估算,以期为不同研究方法所得结果进行比较,为生态系统碳循环过程中植被地下净初级生产力的估算和根系对土壤碳补给研究提供理论依据。
1 研究地点概况[30]本研究在中国科学院海北高寒草甸生态系统定位站进行(37°29′—37°45′ N,101°12′—101°33′ E),该站位于青藏高原的东北隅,平均海拔3200 m,具有明显的高原大陆性气候特点。多年平均气温为-1.7 ℃,多年平均降水量约600 mm,无四季之分,仅有冷暖二季之别,年内无绝对无霜期,相对无霜期约为20 d。研究草地类型为矮嵩草(Kobresia humilis)草甸,群落总覆盖度为60%—90%,以矮嵩草为建群种,次优势种主要有异针茅(Stipa aliena)、羊茅(Festuca ovina)、垂穗披碱草(Elymus nutans)等,伴生种主要有草地早熟禾(Poa pratensis)、高山嵩草(K. pygmaea)、天蓝苜蓿(Medicago lupulina)、黄芪(Astragalus spp.)、麻花艽(Gentiana straminea)、美丽凤毛菊(Saussurea pulchra)、雪白萎陵菜(Potentilla nivea)、 棘豆(Oxytropis spp.)、瑞苓草(Saussurea nigrescens)等。土壤为亚高山草甸土,因受低温环境影响,土壤发育年轻,土层浅薄,有机质含量丰富[31]。
2 研究方法 2.1 根钻法2008年5月,在海北高寒矮嵩草草甸试验地布设3个6 m×12 m的样地。于2008年5—9月,2009年5—9月每月中下旬进行根系采样,每次采样分别在3个样地内随机选择6个样点(2008年每次取18个样品)或4个样点(2009年每次取12个样品)。每个样点用内径5 cm的不锈钢钻由上至下分0—10 cm、10—20 cm二层钻取土芯样品。将取出的土芯样放入自封袋内,编号后带回实验室。在实验室用10目筛网筛去大部分土壤之后,在流水中洗去附着于根系上的土壤。然后将洗净的根系带回实验室,根据外形、颜色、弹性等特征区别死根和活根[32]。将分拣后的根系样品(包括活根和死根)分别装入信封,放入烘箱在65℃烘干至恒重(约48 h),并称重后换算成单位面积上10 cm土层内的活根和死根的生物量(g/m2)。
2.2 内生长袋法2007年7月底,在根钻法取样样地的就近处,布设3个6 m×12 m的样地,以1 m为间隔距离在每个样地中设置72个样点,每个样点用内径5 cm的根钻分两层(每层10 cm)打孔。与此同时,用环刀法测定0—10 cm和10—20 cm的原土容重。然后将事先制作好的生长袋放入洞中(生长袋用网眼2 mm的尼龙网缝制成外径5.2 cm,长24 cm的圆柱形袋子),按上下两层的原土容重及无根土壤的含水量称量所需土壤质量填入生长袋内,生长袋口露出地面约4 cm以标记位置,便于次年和第3年采样时识别。
于2008年5—9月,2009年5—9月每月中下旬对内生长袋内的根系进行采样,基本与连续根钻法取样同一天,每次采样在3个样地内分别随机选取6个样点和4个样点(2008年每次共采18个样品,2009年每次共采12个样品),取出2007年埋设的生长袋。将装有20 cm长土柱的生长袋放入自封袋内,编号后带回实验室。在实验室先将生长袋沿袋壁竖直剪开,用尺量取上下各10 cm两层,在分界处切开,分别用10目筛网筛去大部分土壤,得到0—10 cm和10—20 cm两个土层的根系样品,后续处理同根钻法。
2.3 微根管法2008年8月,在上述两种方法的样地内随机选取4个点,相互间距不小于3 m,安装4个由PVC材料制成的透明圆形管(微根管),微根管内径为5.5 cm,长度为90 cm。首先用特制的钢钻,以与地面成45°的夹角,钻一个与微根管直径相近的管洞,然后将微根管斜插入土壤,插入长度约为70—75 cm,换算成垂直深度达50—55 cm。露出地面的微根管用锡箔纸包裹,以防光线透入,然后用海绵胶带缠绕,在一定程度上保护微根管壁不因撞击而受损;外面再缠绕一层锡箔纸,之后用不透明胶布覆盖,顶端加盖密封,盖长约15 cm,盖子用白色PVC材料制成,防止吸热。微根管自盖子下部至地面的裸露部分用枯落物覆盖。
于2009年5—9月对根系进行连续取样。每隔2周用微根管袖珍影像成像系统(BTC-2 Minirhizotron Camera System; Bartz Technology,Santa Barbara,CA,USA)对管内同一位置的根系生长过程进行影像收集,每次的影像收集工作在1—2 d内完成。图片实际的大小为1.4 cm×1.8 cm(宽×长),放大15倍,每管收集40—45张图片。采集的影像带回实验室用Win RHIZO MF(Regent Instruments Canada Incorporation,Canada)软件进行分析处理,测定每幅影像中根系的直径、长度等数据,并区分根系状态,如活根和死根,根系消失与否等。将白色根和棕色根定义为活根,黑色的根、皮层脱落或表皮褶皱的根定义为死根;另外,如果白色和棕色根系在下一期影像中突然消失,也定义为死根[16, 33, 34]。根系寿命定义为自第一次在影像中出现时刻起到第一次被定义为死亡或消失的时间。
2.4 数据处理方法 2.4.1 根钻法和内生长袋法采用的估算方法对于根钻法和内生长袋法,分别采用了以下3种方法计算地下净初级生产力(BNPP)。
1 ) 极差法 将季节动态中的月最高生物量(Bmax)和最低生物量(Bmin)相减所得,即

2 ) 积分法 将各次测定的细根生物量的增长值(ΔB)累加,即

3 ) 矩阵法[35] 计算方法如表 1所示。
|
死根生物量变化
Change of dead root biomass | 活根生物量变化 Change of live root biomass | ||
| 增加Increase | 减少Decrease | ||
| ΔBdead>ΔBlive | ΔBlive>ΔBdead | ||
| ΔBlive和ΔBdead为相邻两次采样间隔内活根和死根的增加量或减少量;Δ指相邻两次采样得到的根系生物量之差的绝对值; 将每次计算得到的BNPP相加得该年的BNPP(kg hm-2 a-1) | |||
| 增加Increase | BNPP=ΔBlive+ΔBdead | BNPP=ΔBlive+ΔBdead | BNPP=0 |
| 减少Decrease | BNPP=ΔBlive | BNPP=0 | |
根据如下公式计算根系周转速率T(a-1):
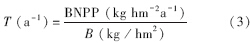
式中,B(kg/hm2)为年平均或年内最大生物量。
2.4.2 微根管法采用的估算方法为了与根钻法和内生长袋法比较基准一致,微根管法采用了0—20 cm土层的观测数据进行根系周转速率的计算,并采用以下两种方法对其进行估算。
首先根据微根管观测窗内得到的根系长度,利用如下公式计算单位土体的根长密度(RLD,mm/cm3):
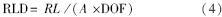
式中,RL(mm)为观测窗中观测到的细根根长;A(cm2)为观测窗面积;DOF(cm)为微根管到周围土壤的距离。DOF一般在0.2—0.3 cm[36, 37, 38, 39]。由于本研究中的草地群落的细根直径较小,计算中DOF取0.2 cm。
然后,通过比根长(SRL,cm/g)将RLD转化为单位体积的生物量(RBD,g/cm3):
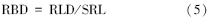
式中,SRL为同期根钻法所得该草甸植被细根(<1 mm)比根长,31.76 cm/g。
最后,通过RBD(g/cm3)乘以取样土壤剖面深度(D,cm)将RBD转化为单位面积根系生物量RBDA(g/cm2):
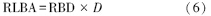
式中,D取最底部的第20个观察窗所处土壤剖面深度,19.8 cm。
利用2.4.1中公式(2)计算BNPP,用ΔRLBA代替公式(2)中的ΔB,ΔRLBA为微根管法相邻两次观测期之间细根净增长量,然后根据公式(3)计算根系周转速率T。
此外,利用统计软件SPSS16.0中的Kaplan-Meier方法进行生存分析,计算细根的中位值寿命(即存活率达到50%时所用的时间,Median root longevity,MRL)。Kaplan-Meier法,又称乘积极限法(简称PL法),由Kaplan-Meier在1958年[40]提出,该方法属于非参数估计法,是利用tk时刻之前各时点上生存概率的乘积来估计在tk时刻的生存率,不需要对被估计的资料分布作任何假设。
3 研究结果 3.1 根钻法估算细根年周转速率表 2给出了采用根钻法观测得到的根系生物量月际动态。从中看出,2008年和2009年0—20 cm土层中5—9月根系生物量平均值分别为(1995 ± 307) g/m2和(1594±281) g/m2;根系生物量最低值分别为(1326 ± 188) g/m2和(1323 ± 256) g/m2,最高值分别为(2476 ± 594) g/m2和(2062 ± 234) g/m2。采用极差法、积分法和矩阵法计算得到2009年地下净初级生产力分别为(7390 ± 281) kg hm-2 a-1、(7390 ± 584) kg hm-2 a-1和(6003 ± 464) kg hm-2 a-1。由于极差法和积分法所估算的BNPP相等,所以根据极差法/(平均生物量或最大生物量)和积分法/(平均生物量或最大生物量)计算所得的根系周转速率也相等,分别为0.46 a-1和0.36 a-1,各种计算方法中用矩阵法/最大生物量所得的根系周转速率值最小。总的来说,根钻法估算得到该草甸细根年周转速率为0.29—0.46 a-1(表 3)。
|
月份
Month | 年份
Year | 根钻法/(g/m2)
Sequential coring method | 内生长袋法/(g/m2)
Ingrowth core method | 微根管法/mm
Minirhizotron method | |||
| 0—10 cm | 10—20 cm | 0—10 cm | 10—20 cm | 0—10 cm | 10—20 cm | ||
| 微根管法对应的值为观测窗内的根长 | |||||||
| 5 | 2008 | 1537±316 | 175±118 | 63±16 | 28±9 | ||
| 2009 | 1332±383 | 93±30 | 180±38 | 79±17 | 97±67 | 35±24 | |
| 6 | 2008 | 1186±205 | 140±40 | 80±27 | 37±15 | ||
| 2009 | 1206±278 | 116±19 | 230±112 | 78±33 | 200±100 | 85±38 | |
| 7 | 2008 | 1879±224 | 204±66 | 137±36 | 50±14 | ||
| 2009 | 1457±263 | 94±24 | 173±35 | 74±22 | 271±148 | 96±61 | |
| 8 | 2008 | 2199±264 | 181±47 | 117±19 | 54±13 | ||
| 2009 | 1481±330 | 131±48 | 286±54 | 97±56 | 244±95 | 89±20 | |
| 9 | 2008 | 2240±652 | 235±45 | 165±33 | 62±19 | ||
| 2009 | 1856±254 | 206±55 | 273±37 | 98±21 | 380±156 | 237±82 | |
|
计算方法
Calculation methods | 周转速率 Turnover rate /a-1 | ||
|
根钻法
Sequential coring method | 内生长袋法
Ingrowth core method | 微根管法
Minirhizotron method | |
|
极差法/(平均生物量或最大生物量)
Max-min method/(mean biomass or maximum biomass) | 0.46或0.36 | 0.43或0.35 | |
|
积分法/(平均生物量或最大生物量)
Integral method/(mean biomass or maximum biomass) | 0.46或0.36 | 0.59或0.48 | 2.99或1.68 |
|
矩阵法/(平均生物量或最大生物量)
Decision matrix method/(mean biomass or maximum biomass) | 0.38或0.29 | 0.63或0.52 | |
|
Kaplan-Meier方法
Kaplan-Meier method | 3.41 | ||
采用内生长袋法观测得到2008年和2009年生长期的5月—9月根系平均生物量分别为(159 ± 23) g/m2和(314 ± 48) g/m2;期间出现的最低生物量分别为(92 ± 16) g/m2和(247 ± 31) g/m2,最高生物量分别为(227 ± 30) g/m2和(383 ± 55) g/m2。采用极差法、积分法和矩阵法估算得到2009年地下净初级生产力分别为(1358 ± 164) kg hm-2 a-1、(1850 ± 308) kg hm-2 a-1和(1987 ± 427) kg hm-2 a-1;进一步计算显示,采用极差法/最大生物量所得的根系年周转速率值最小,为0.35 a-1,用矩阵法/平均生物量所得的根系年周转速率值最大,为0.63 a-1,其余计算方法所得的结果介于0.35—0.63 a-1间(表 3)。
3.3 微根管法估算细根年周转速率表 2中给出了采用微根管法在0—10 cm、10—20 cm土层中观测到根长的月际动态,根据公式(4)将其转化为单位体积土壤中的根系长度,得到整个观测期内根系的平均RLD为(34.40 ± 9.13) mm/cm3,最低值为(13.10 ± 5.46) mm/cm3,最高值为(61.21 ± 12.5) mm/cm3。根据公式(5)和(6)将单位体积的根长密度转化为单位面积的根系生物量后,再根据公式(2)计算得到地下净初级生产力为(3208 ± 587) kg hm-2 a-1,由此计算出的细根年周转速率为1.50 a-1(平均生物量作为分母)或0.84 a-1(最大生物量作为分母)(表 3)。采用Kaplan-Meier方法统计得到微根管法观测的根系中值寿命为107 d,将其换算为细根周转速率,即为3.41 a-1。
4 讨论与结语根钻法是最早被用来测定根系生产力和死亡率的一种方法,也是目前研究根系生物量、生产力和周转最常用的方法之一[41]。内生长袋法是随后发展起来的,它根据生长袋内根系的生物量和埋设生长袋的时间,估算该时段内根系的生产力。在操作上,虽然根钻法可以在研究开展的当年直接取样获得根系生物量,而内生长袋法需要提前埋设生长袋和无根土壤,经过一段时间的稳定期才能开始取样,但从生长袋内采集的根系更容易被分拣成活根和死根。微根管法早在1937年就被提出并应用于根系生长研究[42],它是一种非破坏性的野外观察根系动态的方法[43]。其最大的优点在于可在不干扰根系生长过程的前提下,多次监测单个根系从出生到死亡的全过程,也能记录根系的生长、生产和物候等特征,是当前根系生产力及其周转方面研究中被认为最有前途的方法之一[44]。
本研究同时采用了上述3种观测方法,每种方法分别采用了不同的周转速率计算方法。根钻法和内生长袋法估算细根周转都是基于BNPP的估测数值,而采用不同方法计算得到的BNPP相差很大,导致细根周转的估算结果也表现出较大的差异(表 3),所以BNPP的准确性会直接影响细根周转速率的估算结果及其误差大小。根钻法和内生长袋法中BNPP的估算都是基于根系生物量的季节动态,结合以往文献报道[34, 35, 45, 46, 47, 48]BNPP的具体计算方法主要有:
(1)计算月最大与月最小根系总生物量的差值;
(2)将各次取样测定的根系总生物量间的净增长量分别累加;
(3)将各次取样测定的活根系和死根系间的净增长量累加;
(4)将各次取样测定的活根系间的净增长量累加;
(5)将各次取样测定的植物地下组织各成分(包括根系和根茎)的活生物量净增长量累加。
(6)矩阵法[34],该方法考虑了多种误差因素,并针对性地进行了修正,因而能够更为准确地估算BNPP。
在(2)—(5)方法中,对净增长量累加时,有全部累加的,也有只在相邻两次取样的生物量差异显著时才累加的。Neill[45]的研究显示若是净增长量全部累加,则方法(1)和(2)得到的估算值相近;若只在差异显著时才累加,则估算值比方法(1)和(2)都低;相对于方法(1)和(2),方法(5)计算的结果偏高;方法(3)的计算结果是方法(2)的3倍,并且变异幅度也较大。Pucheta等[46]的比较研究看到,方法(2)—(5)的估算结果之间相差1.74倍,从高到低依次为方法(5)、(2)、(3)和(4)。方法(6)的估计值一般会高于(1)法的估计值[35, 47]。因此是否选择了合适的计算方法对估算结果的准确性有较大影响。如研究生物量季节波动较大的植物根系,不宜选择方法(1)计算;若研究死根比例较高的植物根系,不宜选择方法(3)[45];若采样频度较高,不宜选择方法(2)[48]。根据群落特征和采样频率,本研究采用了上述方法(1)、(2)和(6)3种方法来估算BNPP,结果显示,方法(1)和(2)对根钻法的估算结果相近,与Neill的研究结论吻合[45];但与大多数研究的结果[35, 47]不一致的是,方法(6)的估算值最低,甚至低于方法(1),这可能是因为该草甸植被根系中死根比例较高,从中分拣出死根反而会增加估算误差,因此对矮嵩草高寒草甸方法(2)可能更适合于根钻法中BNPP的计算。对于内生长袋法,方法(1)的计算值最低,方法(6)最高,这与前人总结的结论较为一致[35, 45, 47]。由于内生长袋中的根系是新生长出来的,在相同直径的土柱内其根系量比根钻法少,并且更容易准确分拣出活根和死根,在这种情况下,选择矩阵法估算BNPP是适宜的。
根据文献报道,根系周转速率也有不同的计算方法。如Aber等[49]和Aerts等[50]的研究中采用BNPP/年平均生物量来计算根系周转速率;Hendrick和Pregitzer[33]则用BNPP/最小生物量来计算;Gill和Jackson[17]在综述了当时的研究,发现2/3的研究报道中采用了BNPP/最大生物量方法来计算根系周转速率,说明该方法的使用更为广泛。本研究分别以平均生物量和最大生物量作为分母计算根系周转速率(表 3),比较发现两者的计算结果比较接近,根钻法后者比前者低21.7%—23.7%,内生长袋法中后者比前者低17.5%—18.6%,这表明根系生物量表征方法的选择对根钻法估算根系周转速率的影响更大,内生长袋法相对而言对表征指标的选择敏感性较低,不同指标估算的结果之间一致性较好。
根据上述讨论,将根钻法的BNPP(积分法)/最大生物量的计算结果和内生长袋法的BNPP(矩阵法)/最大生物量的结果进行比较更为合理。此外,用2009年的测定结果进行3种方法的详细比较可能更为可靠。因为对于根钻法,2009年是第2次进行生长季采样工作,实验人员在根系漂洗和分拣等操作上的熟练程度和可靠度比前一年更好,从而可在一定程度上减小人为误差;对于内生长袋法,2009年是生长袋埋设的第2年,袋内的根系更接近于平衡状态,因而降低了因为根系非稳态而导致的系统误差;并且微根管法的计算结果正是来自2009年的观测数据。因此以2009年的结果进行3种方法的比较,不仅可靠性更高,而且还可以避免由于测定时间不匹配导致的气象条件差异的影响。从2009年的研究结果来看,根钻法观测到的细根周转速率为0.36 a-1,内生长袋为0.52 a-1,微根管法为0.84 a-1(积分法)和3.41 a-1(Kaplan-Meier方法)。
众多学者认为,不同观测方法所测定的根系样本的组成不同,可能是导致根系寿命及周转速率测定值不同[13, 25, 51]的重要原因。如Gill[52]发现北美科罗拉多州的矮草干草中的多年生格兰马草(Bouteloua gracilis)的根系中值寿命随直径增粗而延长。Baddeley和Watson[53]研究发现,根系直径每增加0.1 mm,细根死亡率则下降了16%;Wells等[54]对不同直径等级的苹果树细根寿命研究显示,直径为0.1—0.2 mm的细根中值寿命为34 d,0.2—0.3 mm的细根中值寿命为36 d,0.3—0.5 mm的细根中值寿命为97 d,而0.5—1.1 mm的细根中值寿命则大于211 d,即随着细根直径的逐渐增加,其中值寿命也变长。本试验样地的研究结果也表明根系直径与根系寿命呈显著的正相关关系[55]。根钻法是从土芯中收集根系样品,在样品处理过程中,不可避免会丢失大部分最细小的根系,余下的可能是直径相对较粗的根系,因此可能低估了根系的周转速率。内生长袋法采样得到的是直径低于2 mm的细根,样本中根系直径比根钻法得到的根系更细,所以内生长袋内的根系周转较根钻法的快。微根管法虽然能够对细根的生长、衰老、死亡以及分解进行连续地跟踪观察,但由于受到图像采集视野的限制(一般使用的BTC图像采集系统的采集范围为1.8 cm×1.4 cm),通常被观测到的是根系末端直径小、长度短的根,包括绝大部分直径<0.5 mm[32],而这部分根系的寿命也较短,因此微根管法可能高估了根系的周转速率。
尽管根钻法、生长袋法和微根管法都存在一些不足,但依然是目前细根研究中应用最广泛的3种方法。虽然某些不足可通过人为方法克服,但根据作者的研究及以往的研究总结,仍有两方面的原因不可避免地造成各方法测定结果间的巨大差异:(1)同一观测方法因其采用的计算方法不同而导致估算结果产生差异;(2)3种观测方法在采样时所获得的根系样本组成不同。通过本研究的比较分析,认为针对高寒矮嵩草草甸,使用根钻法时,宜采用积分法计算BNPP;使用内生长袋法时,宜采用矩阵法计算BNPP。与根钻法相比,内生长袋法所测得的根系周转速率在不同研究中的可比性更好;而微根管法的估测结果高达内生长袋法的6倍以上。3种观测方法对应的根系样本组成不同,突显出综合应用各观测方法的意义和重要性,从而全面客观地反映细根周转。在今后的研究中,若将根钻法或内生长袋法所得的根系生物量及根系组织中碳氮含量数据,与微根管法所得的根系长度生产量及根系寿命数据相结合研究根系动态,则更能准确估计通过细根周转和分解向土壤系统输送的碳和养分的总量[56]。
| [1] | Santantonio D, Grace J C. Estimating fine-root production and turnover from biomass and decomposition data: a compartmental-flow model. Canadian Journal of Forest Research, 1987, 17(8): 900-908. |
| [2] | Hendrick R L, Pregitzer K S. Patterns of fine root mortality in two sugar maple forests. Nature, 1993, 361(6407): 59-61. |
| [3] | Vogt K A, Crier C C, Vogt D J. Production, turnover, and nutrient dynamics of above-and belowground detritus of world forests. Advances in Ecological Research, 1986, 15: 303-377. |
| [4] | Yang L Y, Li W H. Fine root distribution and turnover in a broad-leaved and Korean pine climax forest of the Changbai Mountain in China. Journal of Beijing Forestry University, 2005, 27(2): 1-5. |
| [5] | Jackson R B, Mooney H A, Schulze E D. A global budget for fine root biomass, surface area, and nutrient contents. Proceedings of the National Academy of Sciences of the United States of America, 1997, 94(14): 7362-7366. |
| [6] | Clark D A, Brown S, Kicklighter D W, Chambers J Q, Thomlinson J R, Ni J, Holland E A. Net primary production in tropical forests: an evaluation and synthesis of existing field data. Ecological Applications, 2001, 11(2): 371-384. |
| [7] | Trumbore S E, Gaudinski J B. The secret lives of roots. Science, 2003, 302(5649): 1344-1345. |
| [8] | Tom M S, Trumbore S E, Chadwick O A, Vitousek P M, Hendricks D M. Mineral control of soil organic carbon storage and turnover. Nature, 1997, 389(6647): 170-173. |
| [9] | He J S, Wang Z Q, Fang J Y. Issues and prospects of belowground ecology with special reference to global climate change. Chinese Science Bulletin, 2004, 49(13): 1226-1234 |
| [10] | Wang W, Peng S S, Fang J Y. Biomass distribution of natural grasslands and its response to climate change in North China. Aric Zone Research, 2008, 25(1): 90-97. |
| [11] | Majdi H, Pregitzer K, Morén A S, Nylund J E, gren G I. Measuring fine root turnover in forest ecosystems. Plant and Soil, 2005, 276(1/2): 1-8. |
| [12] | Milchunas D G. Estimating root production: comparison of 11 methods in shortgrass steppe and review of biases. Ecosystems, 2009, 12(8): 1381-1402. |
| [13] | Tierney G L, Fahey T J. Fine root turnover in a northern hardwood forest: a direct comparison of the radiocarbon and minirhizotron methods. Canadian Journal of Forest Research, 2002, 32(9): 1692-1697. |
| [14] | Robinson D. Scaling the depths: below-ground allocation in plants, forests and biomes. Functional Ecology, 2004, 18(2): 290-295. |
| [15] | Black K E, Harbron C G, Franklin M, Atkinson D, Hooker J E. Differences in root longevity of some tree species. Tree Physiology, 1998, 18(4): 259-264. |
| [16] | Burton A J, Pregitzer K S, Hendrick R L. Relationships between fine root dynamics and nitrogen availability in Michigan northern hardwood forests. Oecologia, 2000, 125(3): 389-399. |
| [17] | Gill R A, Jackson R B. Global patterns of root turnover for terrestrial ecosystems. New Phytologist, 2000, 147(1): 13-31. |
| [18] | Matamala R, Schlesinger W H. Effects of elevated atmospheric CO2 on fine root production and activity in an intact temperate forest ecosystem. Global Change Biology, 2000, 6(8): 967-979. |
| [19] | Matamala R, Gonzalez-Meler M A, Jastrow J D, Norby R J, Schledinger W H. Impacts of fine root turnover on forest NPP and soil C sequestration potential. Science, 2003, 302(5649): 1385-1387. |
| [20] | Norby R J, Ledford J, Reilly C D, Miller N E, O'Neill E G. Fine-root production dominates response of a deciduous forest to atmospheric CO2 enrichment. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(26): 9689-9693. |
| [21] | Zhou B Z, Zhang S G, Fu M Y. Minirhizotron, a new technique for plant root system research: its invention, development and application. Chinese Journal of Ecology, 2007, 26(2): 253-260. |
| [22] | Wang R L, Cheng R M, Xiao W F, Feng X H, Liu Z B, Ge X G, Wang X R, Zhang W Y. Fine root production and turnover in Pinus massoniana plantation in Thress Gorges Reservoir Area of China. Chinese of Journal of Applied Ecology, 2012,23(9): 2346-2352. |
| [23] | Liu J, Yang Q P, Song Q N, Yu D K, Yang G Y, Qing H Y, Shi J M. Strategy of fine root expansion of Phyllostachys pubescens population into evengreen broadleaved forest. Chinese Journal of Plant Ecology, 2013, 37(3): 230-238. |
| [24] | Pei Z Q, Zhou Y, Zheng Y R, Xiao C W. Contribution of fine root turnover to the soil organic carbon cycling in a Reaumuria soongorica community in an arid ecosystem of Xinjiang Uygur Autonomous Region, China. Chinese Journal of Plant Ecology, 2011, 35(11): 1182-1191. |
| [25] | Quan X K, Yu S Q, Shi J W, Yu L Z, Wang Z Q. Minirhizotron and radiocarbon methods: Their application and comparison in estimating fine root longevity. Chinese Journal of Ecology, 2007, 26(3): 428-434. |
| [26] | Huang J X, Lin H, Yang Z J, Lu Z L, Xiong D C, Chen G S, Yang Y S, Xie J S. Estimating fine root production and mortality in subtropical Altingia grlilipes and Castanopsis carlesiiforests. Acta Ecologica Sinica, 2012, 32(14): 4472-4480. |
| [27] | Zhen J X, Huang J X, Wang Z Z, Xiong D C, Yang Z C, Chen G S. Fine root longevity and controlling factors in a Phoebe Bournei plantation. Acta Ecologica Sinica, 2012, 32(23): 7532-7539. |
| [28] | Zhang X Q, Wu K H, Murach D. A review of methods for fine-root production and turnover of trees. Acta Ecologica Sinica, 2000, 20(5): 875-883. |
| [29] | Guo D L. Plant roots: structure. Function, and the role in C and nutrient cycling//Wu J G. Lectures in Modern Ecology (Ⅲ) Advances and Key Topics. Beijing: Higher Education Press, 2007: 92-109. |
| [30] | Zhou X M, Wu Z L. Index of vegetations and plants at Haibei Alpine Meadow Ecosystem Station of Chinese Academy of Science. The Qinghai People Press, 2006: 3-30. |
| [31] | Li Y N, Zhao X Q, Cao G M, Zhao L, Wang Q X. Analyses on climates and vegetation productivity background at Haibei Alpine Meadow Ecosystem Station. Plateau Meteorology, 2004, 23(4): 558-567. |
| [32] | Vogt K A, Persson H. Measuring growth and development of roots.//Lassoie J P, Hinckley T M. Techniques and approaches in forest tree ecophysiology. CRC Press, 1991, 477-501. |
| [33] | Pregitzer K S, Hendrick R L, Fogel R. The demography of fine roots in response to patches of water and nitrogen. New Phytologist, 1993, 125(3): 575-580. |
| [34] | Wells C E, Glenn D M, Eissenstat D M. Changes in the risk of fine-root mortality with age: a case study in peach, Prunus persica (Rosaceae). American Journal of Botany, 2002, 89(1): 79-87. |
| [35] | Fairley R I, Alexander I J. Methods of calculating fine root production in forests//Fitter A H, Atkinson D, Read D J, Usher M B, eds. British Ecological Society Meeting. UK: Biological Sciences, 1985: 37-42. |
| [36] | Itoh S. In situ measurement of rooting density by micro-rhizotron. Soil Science and Plant Nutrition, 1985, 31(4): 653-656. |
| [37] | Sanders J L, Brown D A. A new fiber potic technique for measuring root growth of soybeans under field conditions. Agronomy Journal, 1978, 70(6): 1073-1076. |
| [38] | Taylor H M, Huck M G, Klepper B, Lund Z F. Measurement of soil-grown roots in a rhizotron. Agronomy Journal, 1970, 62(6): 807-809. |
| [39] | Steele S J, Gower S T, Vogt J G, Norman J M. Root mass, net primary production and turnover in aspen, jack pine and black spruce forests in Saskatchewan and Manitoba, Canada. Tree Physiology, 1997, 17(8/9): 577-587. |
| [40] | Kaplan E L, Meier P. Nonparametric estimation from incomplete observations. Journal of the American Statistical Association, 1958, 53(282): 457-481. |
| [41] | Samson B K, Sinclair T R. Soil core and minirhizotron comparison for the determination of root length density. Plant and Soil, 1994, 161(2): 225-232. |
| [42] | Bates G H. A device for the observation of root growth in the soil. Nature, 1937, 139(3527): 966-967. |
| [43] | Johnson M G, Tingey D T, Phillips D L, Storm M J. Advancing fine root research with minirhizotrons. Environmental and Experimental Botany, 2001, 45(3): 263-289. |
| [44] | Hendrick R L, Pregitzer K S. Temporal and depth-related patterns of fine root dynamics in northern hardwood forests. The Journal of Ecology, 1996, 84(2): 167-176. |
| [45] | Neill C. Comparison of soil coring and ingrowth methods for measuring belowground production. Ecology, 1992, 73(5): 1918-1921. |
| [46] | Pucheta E, Bonamici I, Cabido M, Diaz S. Below-ground biomass and productivity of a grazed site and a neighbouring ungrazed exclosure in a grassland in central Argentina. Austral Ecology, 2004, 29(2): 201-208. |
| [47] | McKay H M, Malcolm D C. A comparison of the fine root component of a pure and a mixed coniferous stand. Canadian Journal of Forest Research, 1988, 18(11): 1416-1426. |
| [48] | Vogt K A, Grier C C, Gower S T, Sprugel D G, Vogt D J. Overestimation of net root production: A real or imaginary problem? Ecology, 1986, 67(2): 577-579. |
| [49] | Aber J D, Melillo J M, Nadelhoffer K J, McClaugherty C A, Pastor J. Fine root turnover in forest ecosystems in relation to quantity and form of nitrogen availability: a comparison of two methods. Oecologia, 1985, 66(3): 317-321. |
| [50] | Aerts R, Bakker C, De Caluwe H. Root turnover as determinant of the cycling of C, N, and P in a dry heathland ecosystem. Biogeochemistry, 1992, 15(3): 175-190. |
| [51] | Strand A E, Pritchard S G, McCormack M L, Davis M A, Oren R. Irreconcilable differences: Fine-root life spans and soil carbon persistence. Science, 2008, 319(5862): 456-458. |
| [52] | Gill R A, Burke I C, Lauenroth W K, Milchunas D G. Longevity and turnover of roots in the shortgrass steppe: influence of diameter and depth. Plant Ecology, 2002, 159(2): 241-251. |
| [53] | Baddeley JA, Watson C A. Influences of root diameter, tree age, soil depth and season on fine root survivorship in Prunus avium. Plant and Soil, 2005, 276(1/2): 15-22. |
| [54] | Wells C E. Advances in the Root Demography of Woody Species . USA: The Pennsylvania State University, 1999. |
| [55] | Wu Y B, Deng Y C, Zhang J, Wu J, Tang Y H, Cao G M, Zhang F W, Cui X Y. Root size and soil environments determine root lifespan: evidence from an alpine meadow on the Tibetan plateau. Ecological Research, 2013, 28(3): 493-501. |
| [56] | Majdi H. Root sampling methods-applications and limitations of the minirhizotron technique. Plant and Soil, 1996, 185(2): 255-258. |
| [4] | 杨丽韫, 李文华. 长白山原始阔叶红松林细根分布及其周转的研究. 北京林业大学学报, 2005, 27(2): 1-5. |
| [9] | 贺金生, 王政权, 方精云. 全球变化下的地下生态学: 问题与展望. 科学通报, 2004, 49(13): 1226-123. |
| [10] | 王娓, 彭书时, 方精云. 中国北方天然草地的生物量分配及其对气候的响应. 干旱区研究, 2008, 25(1): 90-97. |
| [21] | 周本智, 张守攻, 傅懋毅. 植物根系研究新技术Minirhizotron的起源、发展和应用. 生态学杂志, 2007, 26(2): 253-260. |
| [22] | 王瑞丽, 程瑞梅, 肖文发, 封晓辉, 刘泽彬, 葛晓改, 王晓荣, 张炜银. 三峡库区马尾松人工林细根生产和周转. 应用生态学报, 2012, 23(9): 2346-2352. |
| [23] | 刘骏, 杨清培, 宋庆妮, 余定坤, 杨光耀, 祁红艳, 施建敏. 毛竹种群向常绿阔叶林扩张的细根策略. 植物生态学报, 2013, 37(3): 230-238. |
| [24] | 裴智琴, 周勇, 郑元润, 肖春旺. 干旱区琵琶柴群落细根周转对土壤有机碳循环的贡献. 植物生态学报, 2011, 35(11): 1182-1191. |
| [25] | 全先奎, 于水强, 史建伟, 于立忠, 王政权. 微根管法和同位素法在细根寿命研究中的应用及比较. 生态学杂志, 2007, 26(3): 428-434. |
| [26] | 黄锦学, 凌华, 杨智杰, 卢正立, 熊德成, 陈光水, 杨玉盛, 谢锦升. 中亚热带细柄阿丁枫和米槠群落细根的生产和死亡动态. 生态学报, 2012, 32(14): 4472-4480. |
| [27] | 郑金兴, 黄锦学, 王珍珍, 熊德成, 杨智成, 陈光水. 闽楠人工林细根寿命及其影响因素. 生态学报, 2012, 32(23): 7532-7539. |
| [28] | 张小全, 吴可红, Dieter Murach. 树木细根生产与周转研究方法评述. 生态学报, 2000, 20(5): 875-883. |
| [29] | 郭大立. 植物根系: 结构、功能及在生态系统物质循环中的地位 // 邬建国. 现代生态学讲座(Ⅲ)学科进展与热点论题. 北京: 高等教育出版社, 2007: 92-109. |
| [30] | 周兴民, 吴珍兰. 中国科学院海北高寒草甸生态系统定位站植被与植物检索表. 青海: 青海人民出版社, 2006: 3-30. |
| [31] | 李英年, 赵新全, 曹广民, 赵亮, 王勤学. 海北高寒草甸生态系统定位站气候、植被生产力背景的分析. 高原气象, 2004, 23(4): 558-567. |