 2014, Vol. 34
2014, Vol. 34文章信息
- 郑挺, 林元烧, 曹文清, 张文静, 郑连明, 王宇杰, 杨位迪
- ZHENG Ting, LIN Yuanshao, CAO Wenqing, ZHANG Wenjing, ZHENG Lianming, WANG Yujie, YANG Weidi
- 北部湾北部生态系统结构与功能——浮游动物空间生态位及其分化
- Ecosystem structure and function in northern Beibu Gulf:zooplankton spatial niche and its differentiation
- 生态学报, 2014, 34(13): 3635-3649
- Acta Ecologica Sinica, 2014, 34(13): 3635-3649
- http://dx.doi.org/10.5846/stxb201211191621
-
文章历史
- 收稿日期:2012-11-19
- 修订日期:2014-2-25






生态位是指在自然生态系统中一个族群在时间、空间上的位置及其与相关种(类)群之间的功能关系[1]。自Grinnell在1917年提出生态位的概念以来,生态位作为生态学研究的核心热点已经持续了将近一个世纪[2, 3]。生态位的研究在理解群落结构功能、群落内物种间关系、生物多样性、群落动态演替和种(类)群进化等方面有重要的作用[4]。目前,国内对于生态位的研究已有不少报道,多集中于陆地植物和动物群落,海洋生物方面的生态位研究工作也有开展,如潮间带[5]、海岛滩涂[6]以及红树林[7]等区域的底栖动物生态位研究,但海洋浮游动物的生态位状况的研究却鲜有报道。本文通过对北部湾北部浮游动物主要优势种的生态位研究,探讨了该区域主要浮游动物的空间分布、生态适应性以及生态相似性,为进一步研究北部湾北部浮游动物种间关系以及关键种的筛选提供理论依据。
1 材料与方法 1.1 研究地区概况北部湾位于我国南海西北部,通过南部湾口及东侧的琼州海峡与南海连通,是一个半封闭浅水海湾。北部湾地处北回归线以南,属亚热带海洋性气候,海域季风盛行,每年11月至次年3月盛行东北风,6—8月盛行偏南风;受海洋暖湿气流影响,具有充沛的水汽来源和水汽输送条件。北部湾温盐分布受到北部湾海水收支、北部湾沿岸的淡水输入和南海高盐水输入的影响以及北部湾环流的作用,总体表现为“夏季北暖南冷,冬季北冷南暖”的特点[8]。海底地势平坦,可分为北部湾北海区、琼州海峡以西以及海南岛以西海区、海南岛南部海区3个部分。湾内叶绿素a浓度存在明显的区域性分布特征,高值区主要位于湾北部雷州半岛以西的近岸海域,而低值区在湾南部远岸深水海域,呈自北向南、由近岸向远岸、由东向西逐渐递减的规律[9]。本文选取北部湾调查海域范围内北纬20°以北的区域(北部湾北部海域)进行着重分析。
1.2 样品采集、处理和分析在北部湾北部海域共设置40个采样站位(图 1),分别于2006年7月(夏季)、2006年12月(冬季)以及2007年4月(春季)和9月(秋季)进行了4次浮游动物采样工作。浮游动物样品采集和室内处理均按《海洋生物生态调查技术规程》进行[10]。根据调查站位的水深,浮游动物样品采用大型浮游生物网(孔径0.505 mm,口径80 cm,网长280 cm;当水深大于30 m时)或浅水Ⅰ型浮游生物网(孔径0.505 mm,口径50 cm,网长145 cm;当水深小于30 m时)由底层至表层垂直拖拽采集样品,样品固定为终浓度5%的中性福尔马林溶液,在实验室内用体视镜鉴定并计数。同船、同时、同站位获得水文和环境化学参数。

|
| 图 1 北部湾北部调查站位示意图 (审图号: GS(2014)659) Fig. 1 Sampling sites in northern Beibu Gulf |
本文所采用的水文、化学和浮游生物等数据均来自于厦门大学海洋与地球学院908项目办公室。
1.3 数据分析 1.3.1 K-优势度曲线K-优势度曲线(K-dominant curve)[11]是依据调查中出现物种的数量的百分比进行降序排列,用图示的方法表示依物种降序排列的物种数量的累积百分比:
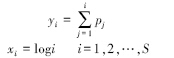
式中,S 是群落中的物种数目,pj为群落中各物种以数量递降顺序排列的第j种的数量百分比,xi为物种降序排列序号的对数,yi为物种丰度的累积百分比。
1.3.2 生态位宽度本文采用较为普遍应用的Levins[12]生态位宽度公式进行计算比较:
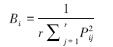
式中,Bi为种i的生态位宽度,取值范围为[0, 1]。Pij为种i在第j个资源状态下的个体数占该种所有个体数的比例;r为资源总数,本研究为采样站位数。
1.3.3 生态位重叠程度本文对种间生态位重叠程度的计算采用Pianka[13]生态位重叠指数:
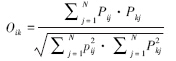
式中,Oik为生态位重叠指数;Pij和Pkj为种i和k在资源j上的优势度。r为资源总数,本研究为采样站位数。
2 结果 2.1 群落K-优势度曲线的季节动态采用K-优势度曲线分析北部湾北部各个季节浮游动物的群落多样性(图 2),春季浮游动物群落多样性要高于冬季且整体趋势与全年的趋势最为接近,冬季又要高于夏季、秋季,夏季和秋季的群落多样性则较为接近。

|
| 图 2 北部湾北部浮游动物各季节和全年的K-优势曲线 Fig. 2 K-dominant curves of zooplankton collected from northern Beibu Gulf in Spring 2007,Summer 2006,Autumn 2007 and Winter 2006,respectively and the whole year |
根据K-优势度曲线排序所得到的种类排序结果,本次调查选取累积百分比80%[14, 15]的优势种(类)群进行生态位分析(表 1)。春季,80%的数量是由20个种(类)群所组成,占总种数的9.35%;夏季,80%的数量是由12个种(类)群所组成,占总种数的4.56%;秋季,80%的数量是由12个种(类)群 组成,占总种数的5.00%;冬季,80%的数量是由16个种(类)群组成,占总种数的7.40%;全年,80%的数量是由20个种(类)群所组成,占总种数的4.27%。
| 编号 | 春季 Spring | 夏季 Summer | 秋季 Autumn | 冬季 Winter | 全年The whole year | |||||
| 种名 Species | % | 种名 Species | % | 种名 Species | % | 种名 Species | % | 种名 Species | % | |
| 1 | 精致真刺水蚤
Euchaeta concinna | 14.63 | 肥胖软箭虫
Sagitta enflata | 18.29 | 肥胖软箭虫
Sagitta enflata | 25.48 | 亚强次真哲水蚤
Subeucalanus subcrassus | 20.10 | 肥胖软箭虫
Sagitta enflata | 18.71 |
| 2 | 肥胖软箭虫
Sagitta enflata | 13.39 | 半索类柱头幼虫
Tornaria larvae | 11.73 | 亚强次真哲水蚤
Subeucalanus subcrassus | 14.94 | 肥胖软箭虫
Sagitta enflata | 14.44 | 亚强次真哲水蚤
Subeucalanus subcrassus | 10.80 |
| 3 | 蛇尾纲长腕幼虫
Ophiopluteus larvae | 6.73 | 短尾类溞状幼体
Brachyura zoea | 10.55 | 汉森莹虾
Lucifer hanseni | 7.89 | 精致真刺水蚤
Euchaeta concinna | 11.39 | 蛇尾纲长腕幼虫
Ophiopluteus larvae | 5.99 |
| 4 | 短尾类溞状幼体
Brachyura zoea | 6.52 | 蛇尾纲长腕幼虫
Ophiopluteus larvae | 8.63 | 小齿海樽
Doliolum denticulatum | 7.24 | 小齿海樽
Doliolum denticulatum | 6.00 | 短尾类溞状幼体
Brachyura zoea | 5.84 |
| 5 | 中型莹虾
Lucifer intermedius | 5.41 | 汉森莹虾
Lucifer hanseni | 8.54 | 蛇尾纲长腕幼虫
Ophiopluteus larvae | 4.92 | 鸟喙尖头溞
Penilia avirostris | 3.55 | 精致真刺水蚤
Euchaeta concinna | 5.55 |
| 6 | 双生水母
Diphyes chamissonis | 3.58 | 拟细浅室水母
Lensia subtiloides | 4.69 | 短尾类溞状幼体
Brachyura zoea | 4.29 | 蛇尾纲长腕幼虫
Ophiopluteus larvae | 3.48 | 汉森莹虾
Lucifer hanseni | 5.26 |
| 7 | 中华哲水蚤
Calanus sinicus | 3.08 | 亚强次真哲水蚤
Subeucalanus subcrassus | 4.47 | 鸟喙尖头溞
Penilia avirostris | 3.91 | 中型莹虾
Lucifer intermedius | 3.13 | 小齿海樽
Doliolum denticulatum | 3.57 |
| 8 | 长尾类幼体
Macrura larvae | 3.07 | 中型莹虾
Lucifer intermedius | 4.36 | 长尾类幼体
Macrura larvae | 2.91 | 双生水母
Diphyes chamissonis | 2.82 | 半索类柱头幼虫
Tornaria larvae | 3.36 |
| 9 | 亚强次真哲水蚤
Subeucalanus subcrassus | 3.00 | 长尾类幼体
Macrura larvae | 3.72 | 椭形长足水蚤
Calanopia elliptica | 2.76 | 软拟海樽
Dolioletta gegenbauri | 2.35 | 中型莹虾
Lucifer intermedius | 3.28 |
| 10 | 鱼卵
Fish eggs | 2.82 | 异摇篮水母
Cunina peregrina | 1.53 | 多毛类幼体
Polychaeta larvae | 1.86 | 微刺哲水蚤
Canthocalanus pauper | 2.32 | 长尾类幼体
Macrura larvae | 3.04 |
| 11 | 长尾住囊虫
Oikopleura longicauda | 2.37 | 球型侧腕水母
Pleurobrachia globosa | 1.41 | 四叶小舌水母
Liriope tetraphylla | 1.73 | 长尾类幼体
Macrura larvae | 2.32 | 鸟喙尖头溞
Penilia avirostris | 2.28 |
| 12 | 多毛类幼体
Polychaeta larvae | 2.28 | 汤氏长足水蚤
Calanopia thompsoni | 1.38 | 长尾类溞状幼体
Macrura zoea | 1.17 | 汉森莹虾
Lucifer hanseni | 2.11 | 拟细浅室水母
Lensia subtiloides | 2.12 |
| 13 | 拟细浅室水母
Lensia subtiloides | 2.12 | 百陶带箭虫
Zonosagitta bedoti | 1.78 | 双生水母
Diphyes chamissonis | 1.79 | ||||
| 14 | 假磷虾幼体
Pseudeuphausia larvae | 2.00 | 短尾类溞状幼体
Brachyura zoea | 1.41 | 多毛类幼体
Polychaeta larvae | 1.54 | ||||
| 15 | 百陶带箭虫
Zonosagitta bedoti | 1.78 | 多毛类幼体
Polychaeta larvae | 1.39 | 椭形长足水蚤
Calanopia elliptica | 1.41 | ||||
| 16 | 针刺真浮萤
Euconchoecia aculeata | 1.75 | 长尾住囊虫
Oikopleura longicauda | 1.20 | 百陶带箭虫
Zonosagitta bedoti | 1.30 | ||||
| 17 | 红住囊虫
Oikopleura rufescens | 1.68 | 微刺哲水蚤
Canthocalanus pauper | 1.01 | ||||||
| 18 | 鸟喙尖头溞
Penilia avirostris | 1.24 | 长尾住囊虫
Oikopleura longicauda | 0.86 | ||||||
| 19 | 半口壮丽水母
Aglaura hemistoma | 1.21 | 鱼卵
Fish eggs | 0.85 | ||||||
| 20 | 锥形宽水蚤
Temora turbinata | 0.95 | 球型侧腕水母
Pleurobrachia globosa | 0.74 | ||||||
| 合计Total | 79.60 | 79.31 | 79.11 | 79.79 | 79.30 | |||||
采用Levins公式计算得到的主要优势种各季节的生态位宽度如图 3所示。春季浮游动物群落中生态位宽度值(Bi)变化范围为0.04—0.62,生态位宽度值最大的3个种(类)群依次为肥胖软箭虫(0.62)、长尾类幼体(0.54)、短尾类溞状幼体(0.49),随后,亚强次真哲水蚤、中型莹虾、针刺真浮萤、精致真刺水蚤和锥形宽水蚤等优势种(类)群的生态位宽度值也较大;夏季浮游动物群落中生态位宽度值变化范围为0.05—0.67,肥胖软箭虫(0.67)和长尾类幼体(0.50)的生态位宽度值仍然很大,而短尾类溞状幼体(0.14)的宽度值锐减,生态位宽度值较大的还有亚强次真哲水蚤、中型莹虾和蛇尾纲长腕幼虫;秋季浮游动物群落中生态位宽度值变化范围为0.05—0.61,生态位宽度值最大的类群为长尾类幼体(0.61),其次为亚强次真哲水蚤(0.57)以及汉森莹虾(0.52),整体来说,秋季主要优势种(类)群之 间生态位宽度值的差异不是十分显著;冬季浮游动物群落中生态位宽度值变化范围为0.03—0.60,百陶带箭虫(0.60)在冬季时的生态位宽度值达到最大,且明显大于该季节出现的其它种(类)群的生态位宽度,与此同时,汉森莹虾(0.05)在该季节的生态位宽度值较秋季时明显变小。在冬季生态位宽度值较大的还有长尾类幼体、亚强次真哲水蚤、短尾类溞状幼体、多毛类幼体和中型莹虾等优势种(类)群。

|
| 图 3 北部湾北部优势浮游动物各季节的生态位宽度 Fig. 3 Niche breadth of dominance zooplankton species collected from northern Beibu Gulf in Spring 2007,Summer 2006,Autumn 2007 and Winter 2006,respectively and the whole year Ml: 长尾类幼体; Se: 肥胖软箭虫; Zb: 百陶带箭虫; Pl: 多毛类幼体; Ss: 亚强次真哲水蚤; Cp: 微刺哲水蚤; Ce: 椭形长足水蚤; Li: 中型莹虾; Dc: 双生水母; Ls: 拟细浅室水母; Ec: 精致真刺水蚤; Ola: 蛇尾纲长腕幼虫; Fe: 鱼卵; Lh: 汉森莹虾; Pg: 球型侧腕水母; Bz: 短尾类溞状幼体; Pa: 鸟喙尖头溞; Tl: 半索类柱头幼虫; Ol: 长尾住囊虫; Dd: 小齿海樽; Ea: 针刺真浮萤; Tt: 锥形宽水蚤; Pla: 假磷虾幼体; Ah: 半口壮丽水母; Cs: 中华哲水蚤; Or: 红住囊虫; Cpe: 异摇篮水母; Ct: 汤氏长足水蚤; Lt: 四叶小舌水母; Mz: 长尾类溞状幼体; Dg: 软拟海樽; I: 广生态位优势种(类)群; II: 中生态位优势种(类)群; III: 窄生态位优势种(类)群(“空白处”表示种(类)群在该季节未进入丰度累积百分比80%,故未予计算) |
对北部湾北部浮游动物全年生态位宽度值进行排序分析,可以分为以下3个类型:(1)广生态位种(类)群,其生态位宽度值0.60—0.77。这一类型的浮游动物可进一步分为两类子类型,一是各个季节均有出现且在大部分的站位点都有较高的个体百分比如长尾类幼体、肥胖软箭虫和亚强次真哲水蚤等,另一子类型则在某些季节大量出现且持续时间长,其全年的生态位宽度仍然很广,如百陶带箭虫、多毛类幼体和微刺哲水蚤等。(2)中生态位种(类)群,生态位宽度值0.30—0.60之间。这一类型的浮游动物也可分为两类子类型,一是如蛇尾纲长腕幼虫、双生水母和球型侧腕水母等类群,其虽然分布广泛但是在每个站位点的个体百分比比重不大或者分布不均匀等特点,另一子类型如精致真刺水蚤、拟细浅室水母、汉森莹虾和鱼卵等则属于某些季节丰度较大的类群。(3)窄生态位种(类)群,生态位宽度值0.08—0.30之间。这一类群的丰度相对较少,并且具有分布不均匀或者只在个别季节出现等特点,如鸟喙尖头溞、半索类柱头幼虫、长尾住囊虫和小齿海樽等。
2.3 生态位重叠北部湾北部海区各季节主要优势种(类)群生态位重叠指数的计测结果见表—表 5。
春季,生态位宽度值较大的种(类)群与其它种(类)群间的生态位重叠较高,亚强次真哲水蚤和百陶带箭虫(0.78)、短尾类溞状幼体和多毛类幼体(0.77)、精致真刺水蚤和亚强次真哲水蚤(0.72)、肥胖软箭虫和短尾类溞状幼体(0.72)、肥胖软箭虫和长尾类幼体(0.71);那些低生态位宽度的种(类)群如长尾住囊虫、鸟喙尖头溞、中华哲水蚤等,往往与其他种(类)群的重叠程度都不高(表 2)。
|
春季Spring
编号No. | Bi | 生态位重叠指数Niche overlap Oik | |||||||||||||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | ||
| 1 : 精致真刺水蚤,2: 肥胖软箭虫,3: 蛇尾纲长腕幼虫,4: 短尾类溞状幼体,5: 中型莹虾,6: 双生水母,7: 中华哲水蚤,8: 长尾类幼体,9: 亚强次真哲水蚤,10: 鱼卵,11: 长尾住囊虫,12: 多毛类幼体,13: 拟细浅室水母,14: 假磷虾幼体,15: 百陶带箭虫,16: 针刺真浮萤,17: 红住囊虫,18: 鸟喙尖头溞,19: 半口壮丽水母,20: 锥形宽水蚤 | |||||||||||||||||||||
| 1 | 0.32 | 1.00 | |||||||||||||||||||
| 2 | 0.62 | 0.45 | 1.00 | ||||||||||||||||||
| 3 | 0.17 | 0.30 | 0.47 | 1.00 | |||||||||||||||||
| 4 | 0.49 | 0.39 | 0.72 | 0.22 | 1.00 | ||||||||||||||||
| 5 | 0.38 | 0.52 | 0.72 | 0.75 | 0.42 | 1.00 | |||||||||||||||
| 6 | 0.25 | 0.26 | 0.45 | 0.40 | 0.30 | 0.55 | 1.00 | ||||||||||||||
| 7 | 0.15 | 0.32 | 0.24 | 0.02 | 0.25 | 0.17 | 0.52 | 1.00 | |||||||||||||
| 8 | 0.54 | 0.37 | 0.71 | 0.30 | 0.66 | 0.61 | 0.40 | 0.27 | 1.00 | ||||||||||||
| 9 | 0.47 | 0.72 | 0.62 | 0.36 | 0.44 | 0.64 | 0.46 | 0.42 | 0.40 | 1.00 | |||||||||||
| 10 | 0.24 | 0.12 | 0.31 | 0.10 | 0.73 | 0.14 | 0.18 | 0.16 | 0.51 | 0.16 | 1.00 | ||||||||||
| 11 | 0.04 | 0.02 | 0.44 | 0.06 | 0.46 | 0.17 | 0.00 | 0.02 | 0.30 | 0.02 | 0.07 | 1.00 | |||||||||
| 12 | 0.31 | 0.37 | 0.67 | 0.30 | 0.77 | 0.46 | 0.29 | 0.19 | 0.41 | 0.58 | 0.49 | 0.20 | 1.00 | ||||||||
| 13 | 0.20 | 0.22 | 0.41 | 0.31 | 0.20 | 0.53 | 0.34 | 0.12 | 0.38 | 0.62 | 0.09 | 0.00 | 0.36 | 1.00 | |||||||
| 14 | 0.21 | 0.47 | 0.17 | 0.05 | 0.29 | 0.14 | 0.13 | 0.16 | 0.19 | 0.25 | 0.22 | 0.01 | 0.18 | 0.09 | 1.00 | ||||||
| 15 | 0.28 | 0.79 | 0.51 | 0.47 | 0.31 | 0.64 | 0.31 | 0.14 | 0.40 | 0.78 | 0.10 | 0.00 | 0.46 | 0.42 | 0.23 | 1.00 | |||||
| 16 | 0.32 | 0.68 | 0.57 | 0.45 | 0.41 | 0.61 | 0.24 | 0.17 | 0.48 | 0.63 | 0.13 | 0.14 | 0.44 | 0.31 | 0.19 | 0.71 | 1.00 | ||||
| 17 | 0.12 | 0.29 | 0.29 | 0.13 | 0.27 | 0.37 | 0.15 | 0.09 | 0.55 | 0.24 | 0.09 | 0.00 | 0.11 | 0.34 | 0.30 | 0.27 | 0.23 | 1.00 | |||
| 18 | 0.05 | 0.01 | 0.36 | 0.06 | 0.18 | 0.11 | 0.00 | 0.01 | 0.20 | 0.10 | 0.06 | 0.35 | 0.11 | 0.01 | 0.00 | 0.02 | 0.05 | 0.01 | 1.00 | ||
| 19 | 0.20 | 0.29 | 0.54 | 0.32 | 0.37 | 0.59 | 0.24 | 0.07 | 0.37 | 0.57 | 0.09 | 0.07 | 0.51 | 0.41 | 0.11 | 0.43 | 0.38 | 0.16 | 0.09 | 1.00 | |
| 20 | 0.32 | 0.47 | 0.59 | 0.30 | 0.49 | 0.53 | 0.45 | 0.34 | 0.54 | 0.55 | 0.19 | 0.10 | 0.47 | 0.31 | 0.16 | 0.40 | 0.43 | 0.17 | 0.08 | 0.48 | 1.00 |
夏季,生态位宽度最大的肥胖软箭虫与中型莹虾、拟细浅室水母、长尾类幼体、蛇尾纲长腕幼虫等种(类)群有着很高的重叠度,分别为0.71,0.68,0.63,0.62。此外,短尾类溞状幼体和长尾类幼体(0.66),蛇尾纲长腕幼虫和异摇篮水母(0.61),汉森莹虾和长尾类幼体(0.65)等种对间也具有较高的生态位重叠度(表 3)。
|
夏季Summer
编号No. | Bi | 生态位重叠指数Niche overlap Oik | |||||||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | ||||
| 1 : 肥胖软箭虫, 2: 半索类柱头幼虫, 3: 短尾类溞状幼体, 4: 蛇尾纲长腕幼虫, 5: 汉森莹虾, 6: 拟细浅室水母, 7: 亚强次真哲水蚤, 8: 中型莹虾, 9: 长尾类幼体, 10: 异摇篮水母, 11: 球型侧腕水母, 12: 汤氏长足水蚤 | |||||||||||||||
| 1 | 0.67 | 1.00 | |||||||||||||
| 2 | 0.10 | 0.31 | 1.00 | ||||||||||||
| 3 | 0.14 | 0.18 | 0.02 | 1.00 | |||||||||||
| 4 | 0.34 | 0.62 | 0.26 | 0.08 | 1.00 | ||||||||||
| 5 | 0.14 | 0.16 | 0.00 | 0.88 | 0.15 | 1.00 | |||||||||
| 6 | 0.27 | 0.68 | 0.09 | 0.09 | 0.51 | 0.21 | 1.00 | ||||||||
| 7 | 0.49 | 0.59 | 0.10 | 0.16 | 0.29 | 0.23 | 0.50 | 1.00 | |||||||
| 8 | 0.35 | 0.71 | 0.11 | 0.09 | 0.43 | 0.00 | 0.28 | 0.33 | 1.00 | ||||||
| 9 | 0.50 | 0.63 | 0.12 | 0.66 | 0.44 | 0.65 | 0.34 | 0.35 | 0.55 | 1.00 | |||||
| 10 | 0.15 | 0.56 | 0.15 | 0.04 | 0.61 | 0.03 | 0.51 | 0.15 | 0.50 | 0.41 | 1.00 | ||||
| 11 | 0.14 | 0.19 | 0.01 | 0.45 | 0.13 | 0.36 | 0.33 | 0.43 | 0.03 | 0.33 | 0.00 | 1.00 | |||
| 12 | 0.05 | 0.05 | 0.00 | 0.53 | 0.04 | 0.30 | 0.04 | 0.15 | 0.03 | 0.22 | 0.00 | 0.46 | 1.00 | ||
秋季,12个主要优势种之间的生态位重叠值大于0.60的高达22对,亚强次真哲水蚤和长尾类幼体(0.77),椭形长足水蚤和四叶小舌(0.78)等一些生态位宽度相近的种(类)群之间表现出较高的生态位重叠程度(表 4)。
|
秋季Summer
编号No. | Bi | 生态位重叠指数Niche overlap Oik | |||||||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | ||||
| 1 : 肥胖软箭虫, 2: 亚强次真哲水蚤, 3: 汉森莹虾, 4: 小齿海樽, 5: 蛇尾纲长腕幼虫, 6: 短尾类溞状幼体, 7: 鸟喙尖头溞, 8: 长尾类幼体, 9: 椭形长足水蚤, 10: 多毛类幼体, 11: 四叶小舌水母, 12:长尾类溞状幼体 | |||||||||||||||
| 1 | 0.51 | 1.00 | |||||||||||||
| 2 | 0.57 | 0.85 | 1.00 | ||||||||||||
| 3 | 0.52 | 0.57 | 0.61 | 1.00 | |||||||||||
| 4 | 0.05 | 0.24 | 0.22 | 0.40 | 1.00 | ||||||||||
| 5 | 0.15 | 0.45 | 0.28 | 0.43 | 0.05 | 1.00 | |||||||||
| 6 | 0.35 | 0.38 | 0.30 | 0.57 | 0.11 | 0.37 | 1.00 | ||||||||
| 7 | 0.19 | 0.51 | 0.50 | 0.58 | 0.82 | 0.21 | 0.16 | 1.00 | |||||||
| 8 | 0.61 | 0.69 | 0.77 | 0.63 | 0.21 | 0.21 | 0.57 | 0.42 | 1.00 | ||||||
| 9 | 0.34 | 0.67 | 0.68 | 0.75 | 0.47 | 0.50 | 0.26 | 0.64 | 0.52 | 1.00 | |||||
| 10 | 0.50 | 0.71 | 0.55 | 0.65 | 0.09 | 0.62 | 0.71 | 0.27 | 0.62 | 0.59 | 1.00 | ||||
| 11 | 0.35 | 0.73 | 0.66 | 0.64 | 0.31 | 0.61 | 0.29 | 0.63 | 0.48 | 0.78 | 0.65 | 1.00 | |||
| 12 | 0.18 | 0.24 | 0.32 | 0.17 | 0.00 | 0.05 | 0.08 | 0.10 | 0.37 | 0.08 | 0.10 | 0.10 | 1.00 | ||
冬季,种(类)群间的生态位重叠程度分布较为均匀,生态位重叠值介于0.30—0.60之间的种对占了全部种对的50.8%,小于0.30的种对占了41.7%。百陶带箭虫作为冬季生态位宽度最大的优势种与其他种(类)群的生态位重叠并不是很高(表 5)。
|
冬季Winter
编号No. | Bi | 生态位重叠指数Niche overlap Oik | |||||||||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | ||
| 1 : 亚强次真哲水蚤, 2: 肥胖软箭虫, 3: 精致真刺水蚤, 4: 小齿海樽, 5: 鸟喙尖头溞, 6: 蛇尾纲长腕幼虫, 7: 中型莹虾, 8: 双生水母, 9: 软拟海樽, 10: 微刺哲水蚤, 11: 长尾类幼体, 12: 汉森莹虾, 13: 百陶带箭虫, 14: 短尾类溞状幼体, 15: 多毛类幼体, 16: 长尾住囊虫 | |||||||||||||||||
| 1 | 0.47 | 1.00 | |||||||||||||||
| 2 | 0.31 | 0.36 | 1.00 | ||||||||||||||
| 3 | 0.33 | 0.50 | 0.12 | 1.00 | |||||||||||||
| 4 | 0.03 | 0.04 | 0.17 | 0.03 | 1.00 | ||||||||||||
| 5 | 0.07 | 0.12 | 0.62 | 0.02 | 0.34 | 1.00 | |||||||||||
| 6 | 0.17 | 0.40 | 0.40 | 0.08 | 0.19 | 0.47 | 1.00 | ||||||||||
| 7 | 0.38 | 0.53 | 0.25 | 0.46 | 0.00 | 0.05 | 0.35 | 1.00 | |||||||||
| 8 | 0.31 | 0.47 | 0.33 | 0.47 | 0.54 | 0.24 | 0.31 | 0.58 | 1.00 | ||||||||
| 9 | 0.10 | 0.19 | 0.58 | 0.05 | 0.06 | 0.87 | 0.39 | 0.13 | 0.26 | 1.00 | |||||||
| 10 | 0.34 | 0.51 | 0.40 | 0.41 | 0.34 | 0.34 | 0.27 | 0.32 | 0.49 | 0.34 | 1.00 | ||||||
| 11 | 0.49 | 0.57 | 0.41 | 0.58 | 0.05 | 0.16 | 0.56 | 0.67 | 0.49 | 0.19 | 0.53 | 1.00 | |||||
| 12 | 0.05 | 0.03 | 0.73 | 0.01 | 0.09 | 0.12 | 0.07 | 0.00 | 0.05 | 0.03 | 0.16 | 0.21 | 1.00 | ||||
| 13 | 0.60 | 0.55 | 0.51 | 0.60 | 0.11 | 0.22 | 0.31 | 0.42 | 0.49 | 0.24 | 0.42 | 0.52 | 0.22 | 1.00 | |||
| 14 | 0.42 | 0.42 | 0.83 | 0.35 | 0.11 | 0.28 | 0.34 | 0.38 | 0.34 | 0.26 | 0.43 | 0.65 | 0.79 | 0.53 | 1.00 | ||
| 15 | 0.40 | 0.52 | 0.50 | 0.20 | 0.50 | 0.36 | 0.39 | 0.39 | 0.68 | 0.41 | 0.53 | 0.41 | 0.12 | 0.45 | 0.41 | 1.00 | |
| 16 | 0.13 | 0.12 | 0.29 | 0.11 | 0.92 | 0.33 | 0.26 | 0.12 | 0.56 | 0.09 | 0.47 | 0.22 | 0.21 | 0.21 | 0.29 | 0.57 | 1.00 |
对北部湾北部全年主要优势种(类)群的生态位重叠值分析结果表明(表 6):20个主要优势种之间生态位重叠值介于0.02—0.89之间,平均生态位重叠指数为0.45,这些种(类)群之间的生态位重叠值大于0.60、属于高度重叠者占29.5%;生态位重叠值介于0.30—0.60之间、属中度重叠者占40.0%;小于0.30的低度重叠者占30.5%。
|
全年Whole
编号No. | Bi | 生态位重叠指数 Niche overlap Oik | |||||||||||||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | ||
| 1 : 肥胖软箭虫, 2: 亚强次真哲水蚤, 3: 蛇尾纲长腕幼虫, 4: 短尾类溞状幼体, 5: 精致真刺水蚤, 6: 汉森莹虾, 7: 小齿海樽, 8: 半索类柱头幼虫, 9: 中型莹虾, 10: 长尾类幼体, 11: 鸟喙尖头溞, 12: 拟细浅室水母, 13: 双生水母, 14: 多毛类幼体, 15: 椭形长足水蚤, 16: 百陶带箭虫, 17: 微刺哲水蚤, 18: 长尾住囊虫, 19: 鱼卵, 20: 球型侧腕水母 | |||||||||||||||||||||
| 1 | 0.75 | 1.00 | |||||||||||||||||||
| 2 | 0.65 | 0.87 | 1.00 | ||||||||||||||||||
| 3 | 0.42 | 0.72 | 0.60 | 1.00 | |||||||||||||||||
| 4 | 0.30 | 0.39 | 0.29 | 0.24 | 1.00 | ||||||||||||||||
| 5 | 0.43 | 0.56 | 0.66 | 0.30 | 0.19 | 1.00 | |||||||||||||||
| 6 | 0.34 | 0.54 | 0.38 | 0.35 | 0.81 | 0.19 | 1.00 | ||||||||||||||
| 7 | 0.08 | 0.36 | 0.47 | 0.24 | 0.09 | 0.09 | 0.30 | 1.00 | |||||||||||||
| 8 | 0.10 | 0.46 | 0.36 | 0.26 | 0.05 | 0.26 | 0.04 | 0.02 | 1.00 | ||||||||||||
| 9 | 0.50 | 0.74 | 0.74 | 0.69 | 0.23 | 0.59 | 0.22 | 0.19 | 0.23 | 1.00 | |||||||||||
| 10 | 0.77 | 0.78 | 0.75 | 0.59 | 0.62 | 0.69 | 0.61 | 0.22 | 0.19 | 0.78 | 1.00 | ||||||||||
| 11 | 0.26 | 0.59 | 0.52 | 0.42 | 0.20 | 0.26 | 0.46 | 0.62 | 0.04 | 0.37 | 0.46 | 1.00 | |||||||||
| 12 | 0.46 | 0.81 | 0.70 | 0.69 | 0.27 | 0.34 | 0.39 | 0.36 | 0.24 | 0.56 | 0.57 | 0.60 | 1.00 | ||||||||
| 13 | 0.49 | 0.73 | 0.74 | 0.53 | 0.18 | 0.56 | 0.18 | 0.21 | 0.39 | 0.75 | 0.63 | 0.31 | 0.51 | 1.00 | |||||||
| 14 | 0.67 | 0.83 | 0.73 | 0.64 | 0.69 | 0.44 | 0.79 | 0.44 | 0.31 | 0.53 | 0.75 | 0.51 | 0.68 | 0.53 | 1.00 | ||||||
| 15 | 0.51 | 0.76 | 0.81 | 0.56 | 0.25 | 0.53 | 0.44 | 0.60 | 0.23 | 0.61 | 0.66 | 0.65 | 0.65 | 0.55 | 0.69 | 1.00 | |||||
| 16 | 0.68 | 0.72 | 0.72 | 0.49 | 0.37 | 0.73 | 0.35 | 0.22 | 0.26 | 0.66 | 0.77 | 0.41 | 0.49 | 0.67 | 0.63 | 0.60 | 1.00 | ||||
| 17 | 0.56 | 0.76 | 0.84 | 0.57 | 0.27 | 0.67 | 0.44 | 0.36 | 0.23 | 0.67 | 0.71 | 0.58 | 0.64 | 0.60 | 0.69 | 0.82 | 0.64 | 1.00 | |||
| 18 | 0.10 | 0.34 | 0.19 | 0.23 | 0.22 | 0.09 | 0.28 | 0.24 | 0.02 | 0.13 | 0.27 | 0.44 | 0.46 | 0.17 | 0.39 | 0.28 | 0.12 | 0.27 | 1.00 | ||
| 19 | 0.40 | 0.46 | 0.33 | 0.30 | 0.89 | 0.23 | 0.74 | 0.08 | 0.12 | 0.27 | 0.63 | 0.22 | 0.31 | 0.27 | 0.64 | 0.29 | 0.46 | 0.32 | 0.17 | 1.00 | |
| 20 | 0.31 | 0.46 | 0.41 | 0.48 | 0.56 | 0.35 | 0.49 | 0.07 | 0.08 | 0.27 | 0.54 | 0.25 | 0.41 | 0.30 | 0.56 | 0.40 | 0.52 | 0.48 | 0.10 | 0.61 | 1.00 |
本次调查各个站位水深范围在12—60m之间,平均水深29m;海域夏季水温最高(30℃左右),冬季最低(20℃左右);调查区域内盐度常年稳定,秋季较其他季节略低;溶解氧浓度在5.74—8.54 mg/L之间,春、冬季较高,夏、秋季较低;叶绿素a浓度在0.46—7.25 mg/m3之间,季节变化趋势由大至小顺序为秋季、冬季、夏季和春季(表 7)。
| 温度/℃
Temperature | 盐度
Salinity | 深度/m
Depth | 溶解氧/(mg/L)
Dissolved oxygen | 叶绿素 a /(mg/m3)
Chlorophyll a | |
| 春季Spring | 21.28±0.92 | 33.27±0.35 | 29.50±12.55 | 7.47±0.32 | 1.67±0.76 |
| 夏季Summer | 30.25±0.32 | 33.15±0.91 | 29.48±12.09 | 6.15±0.13 | 1.89±1.58 |
| 秋季Autumn | 26.24±0.63 | 32.14±0.75 | 29.30±12.20 | 6.62±0.41 | 2.13±1.12 |
| 冬季Winter | 20.30±1.33 | 32.66±0.44 | 29.70±12.70 | 7.69±0.26 | 1.98±1.28 |
选取了与浮游动物分布影响较为密切的温度、盐度、深度、溶解氧和叶绿素a等5个因子作为影响生态位分化的环境因子,采用除趋势典范对应分析(DCCA)来定量分析北部湾北部海域浮游动物种(类)群与各个环境因子的关系(表 8),根据前2个排序轴(横轴为第一排序轴,纵轴为第二排序轴)做排序图(图 4)。物种在排序图上的分布基本能反映其空间分布变化格局。
| 春季Spring | 夏季Spring | 秋季Spring | 冬季Spring | |||||
| Axes 1 | Axes 2 | Axes 1 | Axes 2 | Axes 1 | Axes 2 | Axes 1 | Axes 2 | |
|
物种与环境间的相关系数
Species-environment correlations | 0.885 | 0.819 | 0.790 | 0.518 | 0.876 | 0.706 | 0.827 | 0.626 |
|
物种与环境变化关系的累计比例
Cumulative percentage variance of species-environment relations | 47.7 | 77.5 | 79.1 | 90.9 | 53.7 | 72.3 | 67.2 | 81.3 |
|
特征值总和
Sum of all eigenvalues | 0.770 | 0.679 | 0.410 | 0.704 | ||||
|
典范特征值总和
Sum of all canonical eigenvalues | 0.307 | 0.210 | 0.136 | 0.243 | ||||
|
第一排序轴显著性检验(P)
Test of significance of first canonical axis | 0.002 | 0.002 | 0.002 | 0.002 | ||||
|
所有排序轴显著性检验(P)
Test of significance of all canonical axes | 0.002 | 0.002 | 0.002 | 0.002 | ||||

|
| 图 4 北部湾北部浮游动物DCCA排序图 Fig. 4 DCCA ordination graph of zooplankton species in in northern Beibu Gulf (a)春季物种-环境二维排序: 1 精致真刺水蚤,2 肥胖软箭虫,3 蛇尾纲长腕幼虫,4 短尾类溞状幼体,5 中型莹虾,6 双生水母,7 中华哲水蚤,8 长尾类幼体,9 亚强次真哲水蚤,10 鱼卵,11 长尾住囊虫,12 多毛类幼体,13 拟细浅室水母,14 假磷虾幼体,15 百陶带箭虫,16 针刺真浮萤,17 红住囊虫,18 鸟喙尖头溞,19 半口壮丽水母,20 锥形宽水蚤; (b)夏季物种-环境二维排序:1 肥胖软箭虫,2 半索类柱头幼虫,3 短尾类溞状幼体,4 蛇尾纲长腕幼虫,5 汉森莹虾,6 拟细浅室水母,7 亚强次真哲水蚤,8 中型莹虾,9 长尾类幼体,10 异摇篮水母,11 球型侧腕水母,12 汤氏长足水蚤; (c)秋季物种-环境二维排序: 1 肥胖软箭虫,2 亚强次真哲水蚤,3 汉森莹虾,4 小齿海樽,5 蛇尾纲长腕幼虫,6 短尾类溞状幼体,7 鸟喙尖头溞,8 长尾类幼体,9 椭形长足水蚤,10 多毛类幼体,11 四叶小舌水母,12 长尾类溞状幼体; (d)冬季物种-环境二维排序: 1 亚强次真哲水蚤,2 肥胖软箭虫,3 精致真刺水蚤,4 小齿海樽,5 鸟喙尖头溞,6 蛇尾纲长腕幼虫,7 中型莹虾,8 双生水母,9 软拟海樽,10 微刺哲水蚤,11 长尾类幼体,12 汉森莹虾,13 百陶带箭虫,14 短尾类溞状幼体,15 多毛类幼体,16 长尾住囊虫 |
在春季的排序中,第一排序轴解释了物种数据总差异的47.7%,物种与环境因子的相关系数为0.885,第二排序轴进一步解释了物种数据总差异的29.8%,物种与环境因子的相关系数为0.819,即第一排序轴和第二排序轴共同解释了77.5%的物种与环境因子之间的关系;同理,夏季,秋季和冬季3个季节的排序中,第一排序轴和第二排序轴分别共同解释了90. 9%,72.3%和81. 3%的物种与环境因子之间的关系。各个季节排序分析的前两轴集中了物种-环境因子关系信息的绝大部分,足以反映海域内浮游动物主要优势种(类)群与环境关系的基本特征。根据蒙特卡罗检验(Monte Carlo test)显示,各季节的第一排序轴和总体排序轴的P值均为0.002,即DCCA分析中的各季节第一排序轴和总体排序轴都有非常显著差异。综上,可以认为DCCA排序结果是可信的。
2.4.3 DCCA排序图分析排序图中箭头所处象限表示不同环境因子与排序轴之间的正负相关性;箭头与排序轴的夹角代表着环境因子与排序轴的相关性大小;箭头连线的长度表示群落分布与该因子相关性的大小,连线越长影响越大;2个箭头之间夹角大小代表这2个因子之间相关性大小,夹角越小,相关性越大[16, 17]。
(1)春季
DCCA分析结果表明,第一排序轴与盐度(-0.8625)相关性最大,与温度(-0.7198)、溶解氧(0.6518)、深度(-0.6409)、叶绿素a(0.4854)这些因子也有较好的相关性,即沿第一轴从左至右,盐度、温度和水深逐渐降低,溶解氧和叶绿素a则会逐渐增高。与第二排序轴相关性较明显的因子为深度(-0.5189)和叶绿素a(0.3553)。沿第二轴从下到上,深度逐渐降低,叶绿素a逐渐降低。
大部分的种(类)群集中分布于环境变量的原点周围(图 4(a)),如肥胖软箭虫、中型莹虾、长尾类幼体和亚强次真哲水蚤等,表明这些浮游动物的分布对环境没有特别的偏好。中华哲水蚤、假磷虾幼体适宜生活在水浅、温度和盐度适中、溶解氧低的环境;红住囊虫的分布受深水、高温和高盐环境的影响较大,长尾住囊虫则更倾向于分布在浅水、低温和低盐环境;鸟喙尖头溞是近岸暖水种(类)群,适宜生活 在具有水浅、低温、低盐、高溶解氧和高叶绿素a特点的环境,在图中与春季其他浮游动物种(类)群之间的距离较远。
(2)夏季
DCCA分析结果表明,与第一排序轴关系最密切的依次是深度(-0.7885)、盐度(-0.6414)和温度(-0.6355),其他环境因子与第一轴的相关系数较小。沿第一轴从左至右,水深、盐度和温度逐渐降低。与第二排序轴相关的是温度(-0.2679)、叶绿素a(-0.2237)和溶解氧(-0.2002)等环境因子,但关系都并不密切。沿第二轴从下到上,盐度、溶解氧和温度逐渐降低。
从图 4(b)中可以看出,夏季时浮游动物的分布受水深、盐度和温度的影响较大,而溶解氧和叶绿素a的影响不是十分显著,并且种(类)群在排序图上的分布较春季分散。沿第一排序轴,排序图左侧的异摇篮水母、半索类柱头幼虫等种(类)群倾向于分布在深水、高温且高盐海域,汉森莹虾、球型侧腕水母和汤氏长足水蚤等位于排序图右侧的种(类)群,适宜生活在浅水、低温、低盐的海域。
(3)秋季
DCCA分析结果表明,第一排序轴与深度(0.8287)关系最密切,同时,第一轴与盐度(0.7300)也有较高的相关性。沿第一轴从左至右,深度和盐度逐渐增高。第二排序轴基本反映了叶绿素a(-0.6070)的变化,与其他环境因子的相关性都小于0.3000。沿第二轴从下到上,叶绿素a逐渐降低。
图 4(c)反映了秋季浮游生物种(类)群和环境因子的关系,排序图上各个种(类)群的分布较分散,主要受水深、盐度和叶绿素a的影响,温度和溶解氧对种(类)群分布的影响相对较小。短尾类溞状幼体、多毛类幼体、汉森莹虾和蛇尾纲长腕幼虫位于排序图左侧,该区域具有水浅和低盐特点,其中,蛇尾纲长腕幼虫的分布还受到叶绿素a的影响。排序图右侧,长尾类溞状幼体与水深、盐度和叶绿素a呈正相关。
(4)冬季
DCCA分析结果表明,第一排序轴与溶解氧(0.5103)呈正相关,与温度(-0.7092)、深度(-0.4690)、叶绿素a(-0.4677)和盐度(-0.4021)呈负相关。沿第一轴从左至右,溶解氧逐渐增高,温度、深度、叶绿素a和盐度逐渐降低。第二排序轴与盐度(-0.3647)和叶绿素a(-0.3037)呈负相关,与其他环境因子的相关性都小于0.3000。沿第二轴从下到上,盐度和叶绿素a逐渐降低。
由图 4(d)可见,水深、温度、盐度、溶解氧和叶绿素a对冬季浮游动物种(类)群分布均有一定的影响。除少数几种分布于排序图的右侧外,大部分种(类)群集中分布在环境变量的原点附近。小齿海樽、鸟喙尖头溞和软拟海樽适宜生活在浅水、低温、低盐、低叶绿素a和高溶解氧环境。汉森莹虾与水深、盐度、温度和叶绿素a呈负相关,与溶解氧呈正相关,其实际主要分布于湾北沿岸海域,该区域在冲淡水范围之内。
3 讨论 3.1 北部湾北部浮游动物时空分布与环境因素的联系生态位是种(类)群对多个环境因子生态适应的一个综合结果,它为一个区域内物种的空间分布提供了简单、可靠的生态解释[18]。海洋生态系统与环境之间的相互作用是基于多重的时间和空间尺度上,相对于海洋中其他的生物群落,浮游动物自主运动能力较弱,具有随波逐流的特点,因此,环境因素对浮游动物的生态分布起着主导作用。北部湾北部地处亚热带,为典型的半封闭海湾,海岸线曲折,陆源营养丰富,水文特性复杂,生活在该海域的浮游动物的分布也呈多样化。广生态位的优势种(类)群如百陶带箭虫、亚强次真哲水蚤和长尾类幼体等对环境因子的变化具有较好的生态适应性,因而在北部湾的分布范围广,在四季中与其他许多种(类)群也表现出较高的生态位重叠程度。窄生态位的优势种(类)群对环境的适应性较差,其分布范围在很大程度上受到环境因素的限制,如本次调查中出现的小齿海樽和软拟海樽的高丰度区大部分位于外海水和沿岸水团的交汇区[19]。王学锋等[20]曾对北部湾海樽类的种类组成及分布的研究发现,受3种水系的影响,海樽类的丰度、分布呈现明显的季节波动,研究这些类群的季节动态对海流和水团等变化具有指示意义。此外,一些生态位宽度值较小的优势种(类)群可能对环境的要求近似或互补而表现出较高的生态位重叠程度,如短尾类溞状幼体和汉森莹虾全年时的重叠度达到0.81,它们在夏季、秋季和冬季的重叠度分别达到0.88、0.57和0.79。在海流和水团交汇的区域,浮游动物丰度和种类数剧变,复杂的水文环境导致浮游动物类群表现出大为不同的生态位特性[3],从DCCA图中可知小齿海樽和鸟喙尖头溞对环境的适应性十分接近,但是两者受季节性水团偏移变化的影响[21, 22],种间重叠度在秋季时高达0.82,而在冬季时却只有0.34。通过对北部湾北部海域浮游动物优势种(类)群全年生态位宽度值和重叠程度(图 3,表 6)的分析,总体来说,北部湾北部海域浮游动物优势种(类)群的生态位宽度差异较大,生境内种(类)群之间利用资源环境的互补性较强,这应与该区域的生境多样化并且各优势种对环境适应能力的差异有关。
研究中还发现种(类)群生态位宽度与季节变化有着密切的联系,能够体现浮游动物丰度随季节的动态变化。以汉森莹虾为例,湾内的汉森莹虾几乎全年均有出现,其中,春季时只出现在少数几个站位且丰度很低,夏季时丰度变大但出现频率仍较低,秋季时丰度和出现频率达到全年最大,冬季时丰度又锐减[23],而汉森莹虾生态位宽度值的变化恰恰符合其四季丰度变化情况。生态位宽度客观反映物种在生境中的分布程度,但它不能反映浮游动物具体的丰度大小。如冬季时精致真刺水蚤和双生水母的生态位宽度分别为0.33、0.31,虽然两者的生态位宽度十分接近,精致真刺水蚤的丰度却远远大于双生水母。
3.2 北部湾北部浮游动物的生态适应性浮游动物群落是由互不完全重叠的各个种(类)群所组成,并且群落中的种(类)群受到食物来源、水深和时间等物理、化学以及生物等因素的影响而分散分布[24]。时空生态位的研究主要针对种(类)群丰度在海域内分布情况,一定程度上反映的是种(类)群之间在某个生态因子联系的相似性。DCCA从环境因子的角度通过排序手段研究种(类)群分布与生态环境的关系,揭示了浮游动物在生境中的生态分化现象,对生态位宽度以及重叠程度计测得到一些不能合理解释的结果做出补充说明,如春季时长尾住囊虫和红住囊虫之间的重叠度为0,两者在深度、温度和盐度3个梯度上分化十分明显,长尾住囊虫是北部湾典型的近岸种[25],红住囊虫所分布的海区具有高温高盐的特点[26],生态适应性的不同导致时空分布的差异从而造成两者间重叠度很低,类似的情况还出现在夏季时球型侧腕水母和异摇篮水母、秋季时长尾类溞状幼体和短尾类溞状幼体、冬季时精致真刺水蚤和鸟喙尖头溞等;此外,百陶带箭虫在冬季DCCA图上与其他浮游动物优势种(类)群有着不同程度上的生态分化,其与邻近分布位置的精致真刺水蚤、短尾类溞状幼体和肥胖软箭虫虽然有着较大的重叠度,也仅为0.60、0.53和0.51,生态位上的分化恰恰解释了百陶带箭虫作为冬季生态位宽度值最大的优势种(类)群,在该季节却与其他浮游动物优势种(类)群的重叠度不高的原因。本研究的DCCA四季图中,春季时,大部分的浮游动物优势种(类)群对环境没有特别的偏好性,分布较为集中,但随着温度的升高,夏季和秋季时浮游动物优势种(类)群的分布发生分散,多数的浮游动物优势种(类)群在深度、温度和盐度3个梯度上生态位分化,而在冬季时,海域内水温降到最低,从排序图上直观的发现一部分浮游动物优势种(类)群明显偏向于分布在冬季低温低盐的区域。因此,由DCCA综合分析得到,水深、温度和盐度对浮游动物时空生态分化有直接影响,叶绿素a和溶解氧这些因素对其分化的影响较次之。叶绿素a和溶解氧的高低与浮游植物的分布密切相关,水体理化因素通过控制浮游植物的生长[27],间接影响浮游动物的动态。
| [1] | Qin L. Statistical Ecology. Beijing: China Forestry Press, 2009. |
| [2] | Grinnell J. The niche-relationships of the California Thrasher. The Auk, 1917, 34(4): 427-433. |
| [3] | Soininen J, Heino J, Lappalainen J, Virtanen R. Expanding the ecological niche approach: Relationships between variability in niche position and species richness. Ecological Complexity, 2011, 8(1): 130-137. |
| [4] | Zhang J T. Quantitative Ecology. Beijing: Science Press, 2004. |
| [5] | Zhao Y Q, Zeng J N, Gao A G, Chen Q Z, Shou L, Liao Y B, Huang Y J. Niche of macrozoobenthos in intertidal zone of Jiaojiang Estuary. Chinese Journal of Applied Ecology, 2009, 20(5): 1176-1183. |
| [6] | Ge B M, Bao Y X, Zheng X, Cheng H Y. The structure of the macrobenthic community and niche analysis at a tidal flat of Linkun Island. Acta Ecologica Sinica, 2005, 25(11): 3037-3034. |
| [7] | Tang Y J, Yu S X, Ke Z J. Three methods of niche analysis on mollusk in mangrove area. Chinese Journal of Ecology, 2006, 25(11): 1442-1448. |
| [8] | Chen Z Z, Hu J Y, Sun Z Y, Zhu J. Comparison of temperature and salinity distributions between summer and winter in Beibu Gulf//Li Y, Hu J Y, eds. Beibu Gulf marine scientific research papers (2nd series). Beijing: Ocean Press, 2009: 77-84. |
| [9] | Wu Y C, Guo F, Huang L F. Seasonal changes and distribution characteristics of chlorophyll a in the Beibu Gulf//Lin Y S, Cai L Z, eds. Beibu Gulf marine scientific research papers (3rd series). Beijing: Ocean Press, 2011: 1-10. |
| [10] | Standardization Administration of China. GB/T 12763.6-2007 specifications for oceanographic (survey Part sixth: marine biological survey). Beijing: Standards Press of China, 2007. |
| [11] | Li S F. Status of fish community in East China Sea using the method of abundance-Biomass comparison (ABC) curve. Journal of Fishery Sciences of China, 2008, 15(1): 136-144. |
| [12] | Levins R. Evolution in changing environments: some theoretical explorations. Princeton: Princeton University Press, 1968. |
| [13] | Pianka E R. The structure of Lizard communities. Annual Review of Ecology Systematics, 1973, 4: 53-74. |
| [14] | Newell R C, Seiderer L J, Hitchcock D R. The impact of dredging works in coastal waters: a review of the sensitivity to disturbance and subsequent recovery of biological resources on the seabed. Oceanography and Marine Biology: an Annual Review, 1998, 36: 127-178. |
| [15] | Gilkinson K D, Gordon D C, Maclsaac K G, McKeown D L, Kenchington E L R, Bourbonnais C, Vass W P. Immediate impacts and recovery trajectories of macrofaunal communities following hydraulic clam dredging on Banquereau, eastern Canada\. ICES Journal of Marine Science, 2005, 62(5): 925-947. |
| [16] | TerBraak C J F. Canoco-an extension of decorana to analyze species-environment relationships. Hydrobiologia, 1989, 184(3): 169-170. |
| [17] | TerBraak C J F. Canonical community ordination part I: Basic theory and linear methods. Ecoscience, 1994, 1(2): 127-140. |
| [18] | Hutchinson G E. Cold Spring Harbor Symposium of Quantitative Biology. Concluding remarks, 1957, 22: 415-427. |
| [19] | Zhang J P, Cao W Q, Zheng L M, Yang W D, Lin Y S. Study on Temporal-spatial Distribution of Thaliacea (Tunicata) and the Correlation of Water Masses in the Eastern Beibu Gulf//Lin Y S, Cai L Z, eds. Beibu Gulf marine scientific research papers (3rd series). Beijing: Ocean Press, 2011: 100-113. |
| [20] | Wang X F, Li C H, Jia X P, Liao X L. Species composition and distribution of Thaliacea (Tunicata) in the Beibu Gulf. Fishery Modernization, 2010, 37(2): 59-63. |
| [21] | Chen B. Preliminary discussion on the formation and properties of the Beibu Gulf stream. Journal of Guangxi Academy of Sciences, 1986, 2(2): 92-95. |
| [22] | Tan G H. Preliminary analysis of hydrologic structure and hydrologic feature in the Sea region of the Beibu Gulf. Transactions of Oceanology and Limnology, 1987, 9(4): 7-15. |
| [23] | Wei J M, Lin Y S, Cao W Q, Zheng L M, Yang W D. The Ecology of Lucifer in Beibu Gulf//Lin Y S, Cai L Z, eds. Beibu Gulf marine scientific research papers (3rd series). Beijing: Ocean Press, 2011: 78-89. |
| [24] | Makarewicz J C, Likens G E. Niche Analysis of a Zooplankton Community. Science, 1975, 190(4218): 1000-1003. |
| [25] | Zheng Z, Li S J, Xu Z Z. Marine planktonic biology. Beijing: Ocean Press, 1984. |
| [26] | Xu Z L, Zhang F Y. Species distribution and diversity of Appendicularia in the East China Sea. Journal of Shanghai Fisheries University, 2006, 15(3): 286-291. |
| [27] | Abrantes N, Antunes S C, Pereira M J, Goncalves F. Seasonal succession of cladocerans and phytoplankton and their interactions in a shallow eutrophic lake (Lake Vela, Portugal). Acta Oecologica, 2006, 29(1): 54-64. |
| [1] | 覃林. 统计生态学. 北京: 中国林业出版社, 2009. |
| [4] | 张金屯. 数量生态学.北京: 科学出版社, 2004. |
| [5] | 赵永强, 曾江宁, 高爱根, 陈全震, 寿鹿, 廖一波, 黄逸君. 椒江口潮间带大型底栖动物的生态位. 应用生态学报, 2009, 20(5): 1176-1183. |
| [6] | 葛宝明, 鲍毅新, 郑祥, 程宏毅. 灵昆岛潮间带大型底栖动物群落结构与生态位分析. 生态学报, 2005, 25(11): 3037-3043. |
| [7] | 唐以杰, 余世孝, 柯芝军. 红树林区软体动物生态位的三种分析方法. 生态学杂志, 2006, 25(11): 1442-1448. |
| [8] | 陈照章, 胡建宇,孙振宇, 朱佳. 北部湾东东部海区夏季和冬季温盐平面分布特征比较 // 李炎, 胡建宇. 北部湾海洋科学研究论文集(第2辑). 北京: 海洋出版社, 2009: 77-84. |
| [9] | 吴易超, 郭丰, 黄凌风. 北部湾叶绿素a含量的分布特征与季节变化// 林元烧, 蔡立哲. 北部湾海洋科学研究论文集(第3辑). 北京: 海洋出版社, 2011: 1-10. |
| [10] | 中国国家标准化管理委员会. GB/T 12763.6—2007海洋调查规范(第6部分: 海洋生物调查). 北京: 中国标准出版社, 2007. |
| [11] | 李圣法. 以数量生物量比较曲线评价东海鱼类群落的状况. 中国水产科学, 2008, 15(1): 136-144. |
| [19] | 张建平, 曹文清, 郑连明, 杨位迪, 林元烧. 北部湾东侧海域海樽类时空分布及其与水团相关性研究 // 林元烧, 蔡立哲. 北部湾海洋科学研究论文集(第3辑). 北京: 海洋出版社, 2011: 100-113. |
| [20] | 王学锋, 李纯厚, 贾晓平, 廖秀丽. 北部湾海樽类的种类组成及分布. 鱼类现代化, 2010, 37(2): 59-63. |
| [21] | 陈波. 北部湾水系形成及其性质的初步探讨. 广西科学院学报, 1986, 2(2): 92-95. |
| [22] | 谭光华. 北部湾海区水文结构及其特征的初步分析. 海洋湖沼通报, 1987, 9(4): 7-15. |
| [23] | 魏静梅, 林元烧, 曹文清, 郑连明, 杨位迪. 北部湾莹虾类生态学研究 // 林元烧, 蔡立哲. 北部湾海洋科学研究论文集(第3辑). 北京: 海洋出版社, 2011: 78-89. |
| [25] | 郑重, 李少菁, 许振祖. 海洋浮游生物学. 北京: 海洋出版社, 1984. |
| [26] | 徐兆礼, 张凤英. 东海有尾类种类分布和多样性. 上海水产大学学报, 2006, 15(3): 286-291. |