 2014, Vol. 34
2014, Vol. 34文章信息
- 缪宁, 周珠丽, 史作民, 冯秋红
- MIAO Ning, Zhou Zhuli, SHI Zuomin, FENG Qiuhong
- 岷江冷杉林皆伐后次生群落结构和物种多样性的演替动态
- Successional dynamics of community structure and species diversity after clear-cutting of faxon fir (Abies faxoniana) forest stands
- 生态学报, 2014, 34(13): 3661-3671
- Acta Ecologica Sinica, 2014, 34(13): 3661-3671
- http://dx.doi.org/10.5846/stxb201211151605
-
文章历史
- 收稿日期:2012-11-15
- 修订日期:2014-2-25






2. 阿坝州川西林业局, 理县 623102;
3. 中国林业科学研究院森林生态环境与保护研究所, 国家林业局森林生态环境重点实验室, 北京 100091;
4. 四川省林业科学研究院, 成都 610081
2. Aba State Western Sichuan Forestry Bureau, Li County 623102, China;
3. Key Laboratory on Forest Ecology and Environmental Sciences of State Forestry Administration, Institute of Forest Ecology, Environment and Protection, Chinese Academy of Forestry, Beijing 100091, China;
4. Sichuan Academy of Forestry, Chengdu 610081, China
一般而言,干扰类型、强度和频度在很大程度上决定着退化植被恢复的速度和方向[1, 2],但由于有原生土壤[2]、保留木[3]、先锋种子的传播[4]等因素的存在,森林群落在采伐后的次生演替初期阶段一般速度较快。但受植物群落类型和环境因素的制约,次生群落生物多样性的恢复动态之间差异较大[5, 6, 7]。影响森林群落恢复过程中物种多样性动态的因素是复杂而多样的,群落类型和小生境的异质性、群落发生历史、干扰强度等,都影响着群落内的物种多样性动态[8, 9]。
岷江冷杉(Abies faxoniana)林是川西亚高山地区暗针叶林的代表性类型,在岷江、白龙江及洮河流域形成广袤的森林,是长江和黄河上游重要的生态屏障[10]。1950—1978年间陆续对以岷江冷杉林为主体的川西亚高山天然林进行了高强度和大规模的采伐,之后采伐量逐年减少,至1998年实施了“天然林保护工程”后停采封育。岷江冷杉原始林被破坏后,通过自然恢复,形成了处于不同恢复阶段的次生群落[11]。现已开展的关于川西亚高山次生群落的研究主要包括森林采伐迹地生态学[12, 13]、次生群落结构及优势种群的空间格局分析[14, 15]、次生群落恢复过程中森林生物量的动态变化[16]和林地水文特征动态变化[17]等,然而,对于川西亚高山天然次生群落结构和物种多样性演替动态的研究却鲜见。为深入认识和了解川西亚高山原始暗针叶林皆伐后次生群落的自然演替过程,本文以该区岷江冷杉林皆伐后自然演替不同阶段的次生群落为对象,研究次生演替过程中群落结构和物种多样性的动态变化,以期为该区森林皆伐后恢复重建技术的选择和可持续经营提供科学参考。
1 研究方法 1.1 研究区概况研究区位于四川省阿坝州理县的米亚罗林区(31°24′—31°55′ N,102°35′—103°4′ E)。该地区属典型的高山峡谷地貌,海拔2200—5500 m,坡度多在30°以上。本区属于青藏高原气候区,受太平洋、印度洋及青藏高原三大气团影响,年降水量为600—1100 mm,年蒸发量为1000—1900 mm,1月均温为-8 ℃,7月均温为12.6 ℃,≥ 10 ℃年积温为1200—1400 ℃。该区成土母岩主要为千枚岩、板岩、白云岩等的残坡积风化物,极易风化,主要土壤类型为山地棕色森林土。米亚罗林区植被垂直成带明显,原生森林分布于海拔2400—4200 m处,植被以亚高山暗针叶林为主,主要优势树种为岷江冷杉[15]。岷江冷杉原始林在不同年代被采伐后,通过自然恢复,客观上形成了处于不同演替阶段的次生群落类型,这为研究岷江冷杉林采伐后的次生演替提供了理想的演替序列。
1.2 研究方法 1.2.1 样地设置和数据采集于2004年7—8月,采用空间代替时间的方法,选择立地条件基本一致、岷江冷杉林皆伐后处于不同演替阶段(10、20、30、40、50a)的天然次生群落作为研究对象。在其中随机选取样地(面积为20 m × 20 m),演替10、20、30、40、50a阶段的次生群落样地的数量分别为:6、7、6、8和7个,共34个样地(表 1)。将20 m × 20 m样 地划分为4个10 m × 10 m的样方,测量样方内胸径≥ 5 cm树木的胸径和高度。在每个20 m × 20 m样方的四个角落和中心位置分别设置5个5 m × 5 m的灌木层样方和5个 1 m × 1 m草本层样方,各185个。调查5 m × 5 m样方内每株灌木的种类、数量、高度、盖度等指标,调查1 m × 1 m样方内草本植物的种类、数量、高度、盖度等指标。考虑到层间植物种类较少,将其以灌木计算。
|
演替阶段
Succession stage/a | 样地数量
Number of plot | 海拔
Altitude /m | 坡向
Aspect | 坡位
Slope location | 坡度
Slope /(°) | 树木密度
Density of trees /(株/hm2) | 胸高断面积
Basal area /(m2/hm2) |
| 10 | 6 | 3490—3590 | 北,西北 | 中上 | 25 | 0 | 0 |
| 20 | 7 | 3390—3490 | 北,西北 | 中上 | 30 | 218±35 | 4.75 |
| 30 | 6 | 3250—3350 | 北,西北 | 中上 | 35 | 1370±98 | 18.08 |
| 40 | 8 | 3120—3220 | 西北 | 中上 | 35 | 1753±107 | 28.55 |
| 50 | 7 | 3080—3180 | 北,西北 | 中上 | 35 | 1164±116 | 32.96 |
森林群落乔木层的胸径分布是反映群落结构稳定状态的重要指标。采用分析分布格局样方法中常用的方差均值比和Morisita指数法[18]来分析不同演替阶段群落中树木的水平分布格局。
(1) 方差均值比
假定以V代表方差,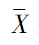 代表平均值,方差/均值比为V/。该比值的含义是,如果V/=1,则个体分布是随机分布;如果V/>1,则个体分布趋于集群分布;若V/<1,则趋于均匀分布。该值的显著性可以用t检验。方差/均值比可以直接计算:
代表平均值,方差/均值比为V/。该比值的含义是,如果V/=1,则个体分布是随机分布;如果V/>1,则个体分布趋于集群分布;若V/<1,则趋于均匀分布。该值的显著性可以用t检验。方差/均值比可以直接计算:
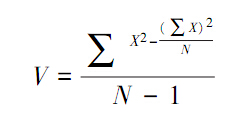
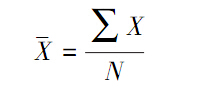
式中,X为样方中的观测值,N是所有样方中观测到的总个体数。t值:
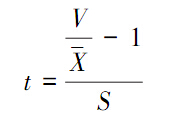
式中,S是标准误差,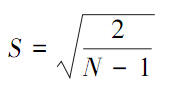
(2) Morisita指数
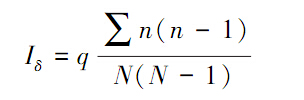
式中,n代表n1,n2,n3,…,nq,分别是q个样方中观测到的个体数;N是所有样方中观测到的总个体数。如果个体是随机分布的,Morisita指数Iδ=1,如果Iδ<1,趋向于均匀分布;若Iδ>1,则为集群分布。该方法的检验可用F检验:
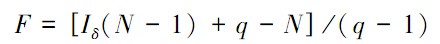
物种多样性作为群落的基本特征,既表征群落的组成结构,也是对环境状况的指示,分别反映了植物群落的不同方面的特点[19],选取的物种多样性测度方法[20]如下:
(1)物种丰富度(S)
S=样方中出现的物种数
(2)Shannon-wiener多样性指数
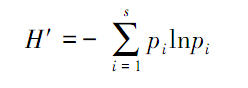
(3)Simpson优势度指数
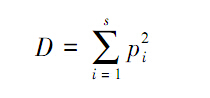
(4)Pielou均匀度指数
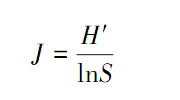
式中,pi为种i的重要值(IV),重要值(IV)的计算式为:
对于乔木

对于灌木和草本

群落中的树木密度在次生演替40年前随着次生演替而不断增大,其中在演替20a到30a阶段树木密度增长迅速。树木密度到演替50a有所下降((1164±116)株/hm2),但群落的胸高断面积随着次生演替而持续增大,达到32.96 m2/hm2(表 1)。经过统计分析,在34个样地中,共记录有107个种,隶属于38科79属。其中有乔木种15个,隶属于8科10属;灌木种30个,隶属于12科21属;草本种62个,隶属于24科49属。不同演替阶段次生群落各层优势种及其重要值变化比较明显,根据优势种组成将该次生群落的演替阶段划分为3个类型:
演替10—20a为悬钩子-蔷薇灌丛,组成群落的主要种类是阳性的灌木和草本,乔木数量极少,为采伐后残留的部分树木,而不是随演替而出现的树木(表 2)。以悬钩子和蔷薇为主的灌木层盖度在60%左右,较密处盖度可达90%。重要值较大的灌木种类主要有黑腺美饰悬钩子(变种)(Rubus subornatus var. melanadenus)、华西蔷薇(Rosa moyesii)、长叶长花忍冬(Lonicera trichosantha var. xerocalyx)、冰川茶藨子(Ribes glaciale)和华西箭竹(Fargesia nitida)(表 3),其中,黑腺美饰悬钩子(变种)、华西蔷薇、长叶长花忍冬、冰川茶藨子等都为典型的阳性灌木。草本植物种类较多且盖度较高,一般在80%左右。重要值较大的草本种类主要有东方草莓(Fragaria orientalis)、拉拉藤(Galium aparine var. echinospermum)、柳兰(原亚种)(Epilobium angustifolium subsp. angustifolium)、大花糙苏(Phlomis megalantha)、糙野青茅(Deyeuxia scabrescens)等阳性草本(表 4)。
演替30a为白桦阔叶林,群落郁闭度在0.6左右,乔木物种较为单一。白桦(Betula platyphylla)重要值最大(55.92),在群落中占据优势地位(表 2)。灌木层盖度在70%左右,重要值排前五位的灌木种类依次为华西蔷薇、长叶长花忍冬、冰川茶藨子、陕甘花楸和柳叶忍冬(Lonicera lanceolata)(表 3)。草本层盖度随林下光照的减弱而继续减小,在70%左右。重要值排前五位的草本种类依次为光柄野青茅 (Deyeuxia levipes)、柳兰(原亚种)(Epilobium angustifolium subsp. angustifolium)、高山耳蕨(Polystichum otophorum)、 千里光(Senecio scandens)和掌叶橐吾(Ligularia przewalskii)(表 4)。在桦木阔叶林演替阶段,先锋种和耐荫种同时存在,但阳性物种的重要值趋于减小,比如黑腺美饰悬钩子(变种),而先锋树种(红桦)、半耐荫(陕甘花楸)和耐荫种(陇塞忍冬和冰川茶藨子)等物种的重要值趋于增大。
|
演替阶段/a
Succession stage | 植物种名
Plant species | 相对多度
Relative abundance | 相对显著度
Relative dominant | 相对频度
Relative frequency | 重要值
Important value |
| 20 | 岷江冷杉Abies faxoniana | 53.13 | 73.32 | 31.25 | 52.56 |
| 四川红杉Larix mastersiana | 14.06 | 5.89 | 12.50 | 10.82 | |
| 白桦Betula platyphylla | 7.81 | 16.84 | 6.25 | 10.30 | |
| 红杉Larix potaninii | 9.38 | 1.82 | 18.75 | 9.98 | |
| 紫果云杉Picea purpurea | 7.81 | 0.89 | 12.50 | 7.07 | |
| 30 | 白桦Betula platyphylla | 85.11 | 55.37 | 27.27 | 55.92 |
| 岷江冷杉Abies faxoniana | 6.69 | 40.25 | 27.27 | 24.74 | |
| 红杉Larix potaninii | 2.74 | 2.87 | 13.64 | 6.41 | |
| 红桦Betula albo-sinensis | 1.52 | 0.35 | 13.64 | 5.17 | |
| 金顶杜鹃Rhododendron faberi | 3.04 | 0.97 | 9.09 | 4.37 | |
| 40 | 红桦Betula albo-sinensis | 51.78 | 40.52 | 15.69 | 36.00 |
| 白桦Betula platyphylla | 14.77 | 21.51 | 15.69 | 17.32 | |
| 岷江冷杉Abies faxoniana | 11.39 | 24.06 | 15.69 | 17.04 | |
| 紫果云杉Picea purpurea | 10.68 | 5.88 | 15.69 | 10.75 | |
| 柳Salix sp. | 2.85 | 2.01 | 9.80 | 4.89 | |
| 50 | 岷江冷杉Abies faxoniana | 17.28 | 42.27 | 11.36 | 23.64 |
| 五尖槭(原亚种)
Acer maximowiczii subsp. maximowiczii | 36.76 | 14.46 | 18.18 | 23.13 | |
| 白桦Betula platyphylla | 12.50 | 25.15 | 11.36 | 16.34 | |
| 红桦Betula albo-sinensis | 12.87 | 10.45 | 18.18 | 13.83 | |
| 西南樱桃Cerasus duclouxiia | 8.09 | 2.68 | 11.36 | 7.38 |
|
演替阶段/a
Succession stage | 植物种名
Plant species | 相对多度
Relative abundance | 相对显著度
Relative dominant | 相对频度
Relative frequency | 重要值
Important value |
| 10 | 黑腺美饰悬钩子(变种)
Rubus subornatus var. melanadenus | 34.27 | 33.88 | 15.56 | 27.90 |
| 华西蔷薇Rosa moyesii | 22.26 | 22.28 | 15.56 | 20.03 | |
| 长叶长花忍冬
Lonicera trichosantha var. xerocalyx | 8.98 | 11.45 | 14.07 | 11.50 | |
| 冰川茶藨子Ribes glaciale | 7.90 | 8.43 | 12.59 | 9.64 | |
| 华西箭竹Fargesia nitida | 10.03 | 6.69 | 7.41 | 8.04 | |
| 20 | 冰川茶藨子Ribes glaciale | 31.74 | 33.83 | 21.26 | 28.94 |
| 长叶长花忍冬
Lonicera trichosantha var. xerocalyx | 18.53 | 12.55 | 17.32 | 16.13 | |
| 陕甘花楸Sorbus koehneana | 8.52 | 19.23 | 17.32 | 15.02 | |
| 黑腺美饰悬钩子(变种)
Rubus subornatus var. melanadenus | 15.76 | 6.87 | 6.30 | 9.64 | |
| 华西蔷薇Rosa moyesii | 7.03 | 9.49 | 11.81 | 9.44 | |
| 30 | 华西蔷薇Rosa moyesii | 39.17 | 36.32 | 21.95 | 32.48 |
| 陇塞忍冬Lonicera tangutica | 16.24 | 15.89 | 21.95 | 18.03 | |
| 冰川茶藨子Ribes glaciale | 13.33 | 12.54 | 15.85 | 13.91 | |
| 陕甘花楸Sorbus koehneana | 9.00 | 8.54 | 12.20 | 9.91 | |
| 柳叶忍冬Lonicera lanceolata | 7.62 | 8.86 | 12.20 | 9.56 | |
| 40 | 华西箭竹Fargesia nitida | 48.99 | 40.58 | 18.32 | 35.96 |
| 鞘柄菝葜Smilax stans | 15.14 | 13.36 | 17.82 | 15.44 | |
| 桦叶荚蒾Vibrnum betulifolium | 12.82 | 14.49 | 13.86 | 13.72 | |
| 冰川茶藨子Ribes glaciale | 4.68 | 4.58 | 10.89 | 6.72 | |
| 紫花卫矛Euonymus porphyreus | 4.14 | 4.30 | 10.89 | 6.44 | |
| 50 | 喜阴悬钩子(原变种)
Rubus mesogaeus var. mesogarus | 27.18 | 24.81 | 12.36 | 21.45 |
| 华西箭竹Fargesia nitida | 27.51 | 17.01 | 11.80 | 18.77 | |
| 鞘柄菝葜Smilax stans | 15.53 | 14.79 | 13.48 | 14.60 | |
| 猕猴桃藤山柳Clematoclethra actinidioides | 3.21 | 13.29 | 10.67 | 9.06 | |
| 冰川茶藨子Ribes glaciale | 5.32 | 5.29 | 10.11 | 6.91 |
演替40—50a为桦木-岷江冷杉针阔混交林,出现了较多的耐荫乔木和灌木种类,强阳性草本种类减少。乔木层郁闭度约为0.7,乔木层优势种为红桦、白桦和岷江冷杉,伴生有山杨(Populus davidiana)、柳(Salix sp.)、紫果云杉(Picea purpurea)等(表 2)。灌木层盖度较高,在80%左右,重要值较高的种主要有华西箭竹、喜阴悬钩子(原变种)(Rubus mesogaeus var. mesogarus)、鞘柄菝葜(Smilax stans)、桦叶荚蒾(Vibrnum betulifolium)、冰川茶藨子(Ribes glaciale)等(表 3)。随次生群落逐渐郁闭,草本层盖度下降较为剧烈,降至30%左右。草本层植物以华北鳞毛蕨(Dryopteris goeringiana)、卵叶韭 (Allium ovalifolium)、高原天名精(Carpesium lipskyi)、 高大鹿药(Smilacina atropurpurea)和大蝎子草(Girardinia diversifolia)等耐荫种占优势,其重要值最大(表 4)。
|
演替阶段/a
Succession stage | 植物种名
Plant species | 相对多度
Relative abundance | 相对显著度
Relative dominant | 相对频度
Relative frequency | 重要值
Important value |
| 10 | 东方草莓Fragaria orientalis | 9.43 | 21.53 | 11.49 | 14.15 |
| 拉拉藤Galium aparine var. echinospermum | 16.21 | 10.37 | 8.11 | 11.56 | |
| 柳兰(原亚种)
Epilobium angustifolium subsp. angustifolium | 17.64 | 8.98 | 5.41 | 10.67 | |
| 大花糙苏Phlomis megalantha | 6.58 | 9.29 | 8.78 | 8.22 | |
| 糙野青茅Deyeuxia scabrescens | 5.73 | 6.81 | 11.49 | 8.01 | |
| 20 | 光柄野青茅Deyeuxia levipes | 13.97 | 31.66 | 13.97 | 19.87 |
| 柳兰
Epilobium angustifolium subsp. angustifolium | 17.65 | 20.02 | 17.65 | 18.44 | |
| 林地早熟禾Poa nemoralis | 8.09 | 14.86 | 8.09 | 10.35 | |
| 六叶葎Galium asperuloides subsp. hoffmeisteri | 0.74 | 0.44 | 0.74 | 0.64 | |
| 高山露珠草Circaea alpina | 0.74 | 0.35 | 0.74 | 0.61 | |
| 30 | 光柄野青茅Deyeuxia levipes | 28.20 | 40.55 | 18.69 | 29.15 |
| 柳兰(原亚种)
Epilobium angustifolium subsp. angustifolium | 29.67 | 27.22 | 17.76 | 24.88 | |
| 高山耳蕨Polystichum otophorum | 6.58 | 3.63 | 14.95 | 8.39 | |
| 千里光Senecio scandens | 9.24 | 5.67 | 5.61 | 6.84 | |
| 掌叶橐吾Ligularia przewalskii | 6.42 | 3.51 | 9.35 | 6.42 | |
| 40 | 华北鳞毛蕨Dryopteris goeringiana | 49.13 | 64.39 | 30.23 | 47.92 |
| 卵叶韭Allium ovalifolium | 9.35 | 4.53 | 15.50 | 9.80 | |
| 高大鹿药Smilacina atropurpurea | 6.68 | 5.07 | 10.08 | 7.27 | |
| 大花糙苏Phlomis megalantha | 3.19 | 7.40 | 6.98 | 5.86 | |
| 大蝎子草Girardinia diversifolia | 5.24 | 3.65 | 7.75 | 5.55 | |
| 50 | 华北鳞毛蕨Dryopteris goeringiana | 26.26 | 29.81 | 24.71 | 26.93 |
| 高原天名精Carpesium lipskyi | 14.69 | 17.58 | 12.94 | 15.07 | |
| 东亚唐松草Thalictrum minus var. hypoleucum | 13.20 | 7.46 | 10.59 | 10.42 | |
| 掌叶橐吾Ligularia przewalskii | 7.27 | 6.80 | 10.59 | 8.22 | |
| 卵叶韭Allium ovalifolium | 9.05 | 3.49 | 11.76 | 8.10 |
演替10a阶段群落为浓密的悬钩子-蔷薇灌丛所覆盖,无乔木树种,其余演替阶段群落乔木层盖度趋于增大,平均胸径不断增长。演替20—50a阶段树木的径级分布主要集中于中、小径级,呈倒“J”型分布(图 1)。
方差均值比和Morisita指数分析结果表明次生群落不同演替阶段的树木均呈显著聚集分布(表 5)。在调查样地中,演替10a阶段尚无胸径≥ 5 cm乔木个体出现,故未纳入计算。随着次生演替,方差均值比的数值趋于增大,说明聚集强度趋于增大,且聚集的显著程度趋于增大(表 5)。Morisita指数所推断出的分布格局都为极显著的聚集分布格局,但聚集强度与演替时间之间无明显的相关性(表 5)。

|
| 图 1 不同演替阶段树木的径级分布 Fig. 1 Diameter class distribution of trees≥ 5 cm DBH at different succession stages |
|
演替阶段/a
Succession stages | 样方数
No. of quadrats | 方差均值比
Variance/ mean value | t | 分布格局
Distribution pattern | Morisita指数
Morisita Index | F | 分布格局
Distribution pattern |
| *表示P<0. 05;**表示P<0. 01 | |||||||
| 10 | 24 | — | — | — | — | — | — |
| 20 | 28 | 1.57 | 2.08* | 聚集分布 | 1.25 | 1.87* * | 聚集分布 |
| 30 | 24 | 1.68 | 2.30* | 聚集分布 | 1.22 | 4.11* * | 聚集分布 |
| 40 | 32 | 2.1 | 4.32* * | 聚集分布 | 1.06 | 2.10* * | 聚集分布 |
| 50年 | 28 | 3.24 | 8.23* * | 聚集分布 | 1.19 | 3.24* * | 聚集分布 |
演替20、30、40、50a阶段的乔木物种丰富度分别为7、7、11和18,在这一过程中群落乔木树种的种类呈增加趋势(图 2)。演替初期的草本物种丰富度最大,随着次生演替,草本物种丰富度趋于减小(图 2),表明群落中的先锋阳性草本逐渐退出群落。而灌木物种丰富度先减少后增多,演替初期最大,在演替30年阶段物种丰富度最小(图 2)。在演替10、20、30、40、50a阶段的群落中,总体物种丰富度分别为54、49、33、54、59,总体物种丰富度先减小,后趋于增大(图 2)。

|
| 图 2 不同演替阶段物种多样性 Fig. 2 Changes of the diversity indices at different succession stages |
随着次生演替,乔木层、灌木层Shannon-Wiener多样性指数趋于增大(图 2),表明乔木层和灌木层的复杂程度趋于增大;草本层的Shannon-Wiener多样性指数趋于减小(图 2),这表明群落中的先锋阳性草本逐渐受到灌木和乔木的遮荫而逐渐退出群落。随着次生演替,灌木层和草本层的Pielou均匀度指数在不同阶段间呈波动的增大趋势,乔木层的Pielou均匀度指数亦不断趋于增大(图 2),这说明随着次生演替,群落各层次中的某些先锋种类独占优势的程度趋于逐渐减弱,物种的分布更趋于均匀。在演替40a阶段前,灌木层和草本层的Simpson优势度指数有逐渐增大的趋势(图 2),但到演替50a阶段却开始减小(图 2),这表明了灌木层和草本层优势度物种先逐渐减少再增加的一个变化;乔木层的Simpson优势度指数在10—20a演替阶段逐渐增大,在演替20a阶段达到最大,此后逐渐减小(图 2),反映了乔木层从桦木等先锋树种大量发生到逐渐衰退,而其他耐荫种逐渐进入群落,导致桦木等先锋树种的优势度趋于减少。
3 讨论岷江冷杉林采伐后所形成的次生群落中,不同演替阶段的树木均呈显著的聚集分布,这反映了自然群落中植物聚集分布的特性,这是因为聚集分布可以增加种内竞争事件,减缓种间的直接竞争[21, 22]。次生群落中的树木密度到演替50a阶段有所下降,但群落的胸高断面积则随着次生演替而持续增大,这反映了该演替系列在演替50a阶段由于树木自疏而导致的密度降低。
本研究中的演替系列中乔木、灌木和草本层的物种组成均表现出耐荫种替代非耐荫种的趋势,这与许多次生演替的研究结果一致[5, 6, 7, 8, 9],这反映了森林正向的次生演替过程中物种组成的变化特点。采伐后森林自然恢复的实质是森林群落的次生演替,是一些物种替代另一些物种的过程[23],每一个阶段系统功能的恢复都会有利于引入构成下一个阶段种类组成中的新种类[24]。川西亚高山暗针叶林在采伐干扰后的恢复演替过程中,群落优势树种红桦和岷江冷杉对光照、空间、养分等的激烈竞争主导了群落的格局及其演替发展方向[15]。然而,森林采伐后的恢复过程,并不仅仅决定于演替各阶段中不同树种的喜光或耐荫性等特性,还要决定于综合的生境条件及其变化特点。
随着恢复演替过程中生物与环境、生物与生物间复杂的相互作用,不同物种间会此消彼长[5],物种多样性在这个过程中会发生显著的变化[25]。本研究中,物种多样性指数的变化趋势与一些研究者的结论一致,比如,在河南宝天曼地区栓皮栎林演替过程中高等植物物种多样性乔木层的丰富度、多样性指数和Simpson优势度指数均趋于增大[5]。在浙江古田山常绿阔叶林演替序列中,乔木层的Shannon-Wiener指数亦呈先增大后变小的趋势[26]。本研究中的物种多样性指数变化趋势与另一些研究则不一致,比如,本研究中总体物种丰富度在悬钩子-蔷薇灌丛到桦木阔叶林阶段中趋于减小,在桦木阔叶林到针阔混交林阶段趋于增大。然而,在缙云山森林次生演替过程中,总体物种丰富度呈先变大后变小的趋势[27]。对于草本植物的Shannon-Wiener多样性指数,本研究中该指数随着演替时间的增加呈减小趋势,这与退化喀斯特森林恢复演替过程中的该指数动态类似[6],然而,在广西大明山中山森林植被恢复过程中该指数则变化较为复杂,未表现出明显的规律性[7]。这些多样性的变化差异反映了在植物群落次生演替的过程中其物种多样性并非仅一种变化趋势而是存在着多种不同的变化趋势。
植被恢复是退化生态系统重建的重要途径,植被恢复过程物种多样性的变化反映了植被的恢复程度[28]。本研究反映了川西亚高山岷江冷杉林采伐后正向演替的一般过程:随着次生演替,从悬钩子-蔷薇灌丛演替至桦木-岷江冷杉针阔混交林,尽管该阶段桦木等阔叶树种仍占优势,但地带性植被优势种岷江冷杉大量出现,意味着次生群落趋于向该地区的原有顶极群落——岷江冷杉林演替[29]。川西亚高山森林采伐后,如果停止人为干扰,由于有原生土壤和保留木(种源)的存在,并依靠自身的反馈系统,是可以自然恢复的。由于受研究时间和空间尺度的制约,本研究揭示的岷江冷杉林皆伐后次生群落的结构和物种多样性动态仅仅是一种演替中期以前的变化趋势,而并非明确的变化规律。要深入理解和揭示采伐后川西亚高山次生群落的演替动态,还需要更大时空尺度的研究。
致谢: 植物标本由四川省林业科学研究院的牟克华先生鉴定;岳永杰、窦源东、陈天学、李玉伟等参与野外调查,特此致谢。
| [1] | Pascarella J B, Aide T M, Serrano M I, Zimmerman J K. Land-use history and forest regeneration in the Cayey mountains, Puerto Rico. Ecosystems, 2000, 3(3): 217-228. |
| [2] | Chazdon R L. Tropical forest recovery: Legacies of human impact and natural disturbances. Perspectives in Plant Ecology, Evolution and Systematics, 2003, 6: 51-71. |
| [3] | Miao N, Liu S R, Shi Z M, Ma J M, Wang H. A review of ecological effects of remnant trees in degraded forest ecosystems after severe disturbances. Acta Ecologica Sinica, 2013, 33(13): 3889-3897. |
| [4] | Holl K D. Factors limiting tropical rain forest regeneration in abandoned pasture: Seed rain, seed germination, microclimate, and soil. Biotropica, 1999, 31(2): 229-242. |
| [5] | Shi Z M, Liu S R, Chen R M. Changes in plant species diversity in a restoration sequence of Quercus variabilis forest stands in Bao Tianman Mountain. Acta Phytoecologica Sinica, 1998, 22(5): 415-421. |
| [6] | Yu L F, Zhu S Q, Ye J Z, Wei L M, Chen Z R. Dynamics of a degraded karst forest in the process of natural restoration. Scientia Silvae Sinicae, 2002, 38(1): 1-7. |
| [7] | Wen Y G, Yuan C A, Li X X, He T P, Lai J Y. Development of species diversity in vegetation restoration process in mid-mountain region of Damingshan, Guangxi. Acta Phytoecologica Sinica, 1998, 22(1): 33-40. |
| [8] | Shen Q, Zhang J, Zhu J R, Jiang B, Ge Y, Liu Q X, Chang J. Changes of species composition and diversity in the restoration processes of ecological public-welfare forests in Zhejiang, East China. Acta Ecologica Sinica, 2005, 25(9): 2131-2138. |
| [9] | Kubota Y, Katsuda K, Kikuzawa K. Secondary succession and effects of clear-logging on diversity in the subtropical forests on Okinawa Island, southern Japan. Biodiversity and Conservation, 2005, 14(4): 879-901. |
| [10] | Liu S R, Shi Z M, Ma J M, Zhao C M, Zhang Y D, Liu X L. Ecological strategies for restoration and reconstruction of degraded natural forests on the upper reaches of the Yangtze River. Scientia Silvae Sinicae, 2009, 45(2): 120-124. |
| [11] | Zhang Y D, Liu S R, Zhao C M. Spatial pattern of sub-alpine forest restoration in west Sichuan. Chinese Journal of Applied Ecology, 2005, 16(9): 1706-1710. |
| [12] | Ma X H. Cutting and soil & water conservation of alpine dark coniferous forest in west Sichuan. Scientia Silvae Sinicae, 1963, 8(2): 149-158. |
| [13] | Shi L X, Wang J X, Xu Y M, Hou G W. Earlier-stage succession of vegetation on the clear-cuts in Mialou forest district in western Sichuan. Acta Phytoecologica et Geobotanica Sinica, 1988, 12(4): 306-313. |
| [14] | Miao N, Shi Z M, Feng Q H, Liu X L, He F. Spatial pattern analysis of Abies faxoniana population in sub-alpine area in western Sichuan. Scientia Silvae Sincae, 2008, 44(12): 1-6. |
| [15] | Miao N, Liu S R, Shi Z M, Yu H, Liu X L. Spatial patterns of dominant tree species in sub-alpine Betula-Abies forest in west Sichuan of China. Chinese Journal of Applied Ecology, 2009, 20(6): 1263-1270. |
| [16] | Zhang Y D, Liu Y C, Liu S R, Zhang X H. Dynamics of stand biomass and volume of the tree layer in forests with different restoration approaches based on tree-ring analysis. Chinese Journal of Plant Ecology, 2012, 36(2): 117-125. |
| [17] | Zhang Y D, Zhao C M, Liu S R. Woodland hydrological effects of spruce plantations and natural secondary series in sub-alpine region of western Sichuan. Journal of Natural Resources, 2004, 19(6): 761-768. |
| [18] | Greig-Smith P. Quantitative Plant Ecology. Oxford: Blackwell, 1983: 54-104. |
| [19] | Brkenhielm S, Liu Q. Long-term effects of clear-felling on vegetation dynamics and species diversity in a boreal pine forest. Biodiversity and Conservation, 1998, 7(2): 207-220. |
| [20] | Magurran A E. Ecological Diversity and Its Measurement. Princeton: Princeton University Press, 1988. |
| [21] | Li B, Chen J K, Watkinson A R. A literature review on plant competition. Chinese Bulletin of Botany, 1998, 15(4): 18-29. |
| [22] | Tokeshi M. Species Coexistence: Ecological and Evolutionary Perspectives. Oxford: Blackwell Publishing, 1999. |
| [23] | Cain M D, Shelton M G. Secondary forest succession following reproduction cutting on the upper coastal plain of southeastern Arkansas, USA. Forest Ecology and Management, 2001, 146(1/3): 223-238. |
| [24] | Zhao P, Peng S L. Species and species diversity in relation with restoration and persistence of degraded ecosystem functions. Chinese Journal of Applied Ecology, 2001, 12(1): 132-136. |
| [25] | Zhu H G, Li Y Q, Wen Y G, Liang H W, Lin J Y, Nong Y. The dynamics of the structure and plant species diversity of evergreen broadleaved forests in Damingshan National Nature Reserve after a severe ice storm damage in 2008, China. Acta Ecologica Sinica, 2011, 31(19): 5571-5577. |
| [26] | Hu Z H, Yu M J. Species diversity in four saccessional communites of evergreen broad-leaved forest in Gutian Mountain, Zhejiang Province. Chinese Journal of Ecology, 2006, 25(6): 603-606. |
| [27] | Guo Q B, Li Y C, Li X G. Dynamics of species diversity in secondary succession series of forest communities in Jinyun Mt. Chinese Journal of Applied Ecology, 1999, 10(5): 521-524. |
| [28] | Wang Y J, Tao J P, Zhang W Y, Zang R G, Wang W, Li Z F, Li Y. Dynamics of species diversity in vegetation restoration on Tudiling of Mao County, southwest China. Acta Ecologica Sinica, 2006, 26(4): 1028-1036. |
| [29] | Ma J M, Liu S R, Shi Z M, Zhang Y D, Miao N. Natural regeneration of Abies faxoniana along restoration gradients of subalpine dark coniferous forest in western Sichuan, China. Chinese Journal of Plant Ecology, 2009, 33(4): 646-657. |
| [3] | 缪宁, 刘世荣, 史作民, 马姜明, 王晖. 强度干扰后退化森林生态系统中保留木的生态效应研究综述. 生态学报, 2013, 33(13): 3889-3897. |
| [5] | 史作民, 刘世荣, 程瑞梅. 宝天曼地区栓皮栎林恢复过程中高等植物物种多样性变化. 植物生态学报, 1998, 22(5): 415-421. |
| [6] | 喻理飞, 朱守谦, 叶镜中, 魏鲁明, 陈正仁. 退化喀斯特森林自然恢复过程中群落动态研究. 林业科学, 2002, 38(1): 1-7. |
| [7] | 温远光, 元昌安, 李信贤, 和太平, 赖家业. 大明山中山植被恢复过程植物物种多样性的变化. 植物生态学报, 1998, 22(1): 33-40. |
| [8] | 沈琪, 张骏, 朱锦茹, 江波, 葛滢, 刘其霞, 常杰. 浙江省生态公益林植被恢复过程中物种组成及多样性的变化. 生态学报, 2005, 25(9): 2131-2138. |
| [10] | 刘世荣, 史作民, 马姜明, 赵常明, 张远东, 刘兴良. 长江上游退化天然林恢复重建的生态对策. 林业科学, 2009, 45(2): 120-124. |
| [11] | 张远东, 刘世荣, 赵常明. 川西亚高山森林恢复的空间格局分析. 应用生态学报, 2005, 16(9): 1706-1710. |
| [12] | 马雪华. 川西高山暗针叶林区的采伐与水土保持. 林业科学, 1963, 8(2): 149-158. |
| [13] | 史立新, 王金锡, 宿以民, 侯广维. 川西米亚罗地区暗针叶林采伐迹地早期植被演替过程的研究. 植物生态学与地植物学学报, 1988, 12(4): 306-313. |
| [14] | 缪宁, 史作民, 冯秋红, 刘兴良, 何飞. 川西亚高山岷江冷杉种群的空间格局分析. 林业科学, 2008, 44(12): 1-6. |
| [15] | 缪宁, 刘世荣, 史作民, 喻泓, 刘兴良. 川西亚高山红桦-岷江冷杉林优势种群的空间格局分析. 应用生态学报, 2009, 20(6): 1263-1270. |
| [16] | 张远东, 刘彦春, 刘世荣, 张笑鹤. 基于年轮分析的不同恢复途径下森林乔木层生物量和蓄积量的动态变化. 植物生态学报, 2012, 36(2): 117-125. |
| [17] | 张远东, 赵常明, 刘世荣. 川西亚高山人工云杉林和自然恢复演替系列的林地水文效应. 自然资源学报, 2004, 19(6): 761-768. |
| [21] | 李博, 陈家宽, 沃金森 A R. 植物竞争研究进展. 植物学通报, 1998, 15(4): 18-29. |
| [24] | 赵平, 彭少麟. 种、种的多样性及退化生态系统功能的恢复和维持研究. 应用生态学报, 2001, 12(1): 132-136. |
| [25] | 朱宏光, 李燕群, 温远光, 梁宏温, 林建勇, 农友. 特大冰冻灾害后大明山常绿阔叶林结构及物种多样性动态. 生态学报, 2011, 31(19): 5571-5577. |
| [26] | 胡正华, 于明坚. 浙江古田山常绿阔叶林演替序列研究:群落物种多样性. 生态学杂志, 2006, 25(06): 603-606. |
| [27] | 郭全邦, 李玉成, 李旭光. 缙云山森林次生演替序列群落的物种多样性动态. 应用生态学报, 1999, 10(5): 521-524. |
| [28] | 王永健, 陶建平, 张炜银, 臧润国, 王微, 李宗峰, 李媛. 茂县土地岭植被恢复过程中物种多样性动态特征. 生态学报, 2006, 26(4): 1028-1036. |
| [29] | 马姜明, 刘世荣, 史作民, 张远东, 缪宁. 川西亚高山暗针叶林恢复过程中岷江冷杉天然更新状况及其影响因子. 植物生态学报, 2009, 33(4): 646-657. |