 2014, Vol. 34
2014, Vol. 34文章信息
- 官文江, 高峰, 李纲, 陈新军
- GUAN Wenjiang, GAO Feng, LI Gang, CHEN Xinjun
- 复合种群管理的风险评估——以日本鲐为例
- Risk evaluation for meta-population management:a case study using chub mackerel
- 生态学报, 2014, 34(13): 3682-3692
- Acta Ecologica Sinica, 2014, 34(13): 3682-3692
- http://dx.doi.org/10.5846/stxb201211121588
-
文章历史
- 收稿日期:2012-11-12
- 修订日期:2014-2-25






2. 大洋渔业资源可持续开发省部共建教育部重点实验室, 上海海洋大学, 上海 201306
2. The Key Laboratory of Sustainable Exploitation of Oceanic Fisheries Resources, Ministry of Education, Shanghai 201306, China
在渔业资源评估与管理中,渔业资源常被简化为空间分布均匀的单一种群[1, 2, 3]。但渔业资源常由多个地方种群或产卵种群组成,种群间既相对独立,又存在个体或繁殖体交流,构成复合种群[4]。研究表明[5],有越来越多的海洋鱼类种群应为复合种群,如大西洋鲱鱼[6]。我国东、黄海日本鲐(Scomber japonicus)也具有复合种群的特点:东、黄海日本鲐由东海西部群与五岛西部群组成[7];从标志放流[8, 9]、形态学比较[10]、仔幼鱼扩散模拟[11]等结果看,两种群存在混合与生殖交流可能[12]。
鱼类种群的空间结构对资源评估与管理的影响已引起了学者的注意[3, 13, 14]。Kerr等[3]认为考虑种群的空间结构有助于提高资源评估与管理的能力。忽略鱼类种群的空间结构,常导致渔业资源的过度开发[15]。但目前,东、黄海日本鲐的资源评估与管理并未考虑其复合种群的特点[16, 17, 18]。为此,本文基于复合种群概念,模拟了东、黄海日本鲐的动态变化,以用于分析在复合种群条件下,日本鲐资源评估及管理方案的风险,为具有复合种群特点的渔业资源的开发与保护提供理论依据。
1 材料与方法 1.1 种群动态模拟为简化,假设东、黄海日本鲐的东海西部群(由SI表示)与五岛西部群(由SII表示)具有明确的空间分布界限,分布面积相等,且按一定比例进行种群交换。为此,其种群动态表示:
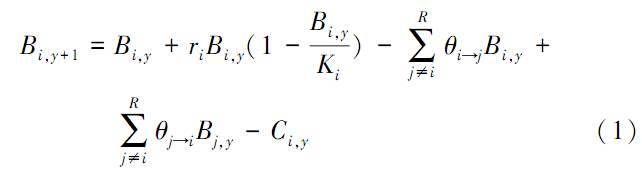
式中,B为资源量,r为内禀增长率,K为环境承载容量,C为捕捞产量,下标y为年份,i,j为种群SI或SII。θi->j为交换矩阵元素,表示种群i迁入到种群j的比例,R为种群集合。
观测的单位捕捞努力量渔获量(CPUE,Catch Per Unit Effort)数据由式(2)计算:
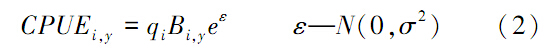
式中,q为捕捞系数,ε为观测误差,假设其服从正态分布,σ可由CPUE变异系数(coefficient of variability,CV)计算:
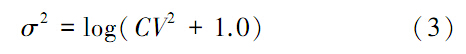
根据交换矩阵、CPUE的变异系数、环境承载容量、内禀增长率的不同,共模拟了12种情况(表 1),其中交换矩阵Θ的设置见表 2。
|
模拟情况
Scenarios | 交换矩阵
Exchange matrix | 变异系数
CV |
环境承载容量
K/104t | 内禀生长率
r | 最大可持续
产量MSY /104t | 最大可持续
产量生物量 BMSY/104t | ||
| SI | SII | SI | SII | |||||
| CV: Coefficient of Variability; K: Carrying capacity; r: Intrinsic growth rate; SI: West East China Sea Population; SII: West Got Islands Population; MSY: Maximum Sustainable Yield; BMSY: Biomass at Maximum Sustainable Yield;Θ交换矩阵exchange matrix | ||||||||
| 1 | Θ | 0.05 | 155 | 155 | 0.88 | 0.88 | 68.2 | 155 |
| 2 | Θ | 0.25 | 155 | 155 | 0.88 | 0.88 | 68.2 | 155 |
| 3 | Θ×2.5 | 0.05 | 155 | 155 | 0.88 | 0.88 | 68.2 | 155 |
| 4 | Θ×2.5 | 0.25 | 155 | 155 | 0.88 | 0.88 | 68.2 | 155 |
| 5 | Θ | 0.05 | 155 | 120 | 0.88 | 0.88 | 60.5 | 137.5 |
| 6 | Θ | 0.25 | 155 | 120 | 0.88 | 0.88 | 60.5 | 137.5 |
| 7 | Θ×2.5 | 0.05 | 155 | 120 | 0.88 | 0.88 | 60.5 | 137.5 |
| 8 | Θ×2.5 | 0.25 | 155 | 120 | 0.88 | 0.88 | 60.5 | 137.5 |
| 9 | Θ | 0.05 | 155 | 155 | 0.88 | 0.80 | 65.1 | 155 |
| 10 | Θ | 0.25 | 155 | 155 | 0.88 | 0.80 | 65.1 | 155 |
| 11 | Θ×2.5 | 0.05 | 155 | 155 | 0.88 | 0.80 | 65.1 | 155 |
| 12 | Θ×2.5 | 0.25 | 155 | 155 | 0.88 | 0.80 | 65.1 | 155 |
模拟中,中国东、黄海沿海8省市(天津、河北、辽宁、江苏、山东、上海、浙江、福建)鲐鱼产量数据来自1997—2010年中国渔业统计年鉴, 并设为种群SI的捕捞产量,日本、韩国的日本鲐产量数据[18]设为种群SII的捕捞产量。此外,1997年种群SI的生物量(BSI,1997)与1997年种群SII的生物量(BSII,1997)分别设为80×104t与73×104t[18],qI与qII分别为0.00675与0.0129。每种情况的一次模拟,共获得两亚种群14a(1997—2010年)的CPUE数据,每种情况重复模拟2000次,获得2000组数据用于分析。
| 移出From | 移入To | |
| 种群SI Subpopulation SI | 种群SII Subpopulation SII | |
| 种群SI Subpopulation SI | — | 0.08 |
| 种群SII Sub-population SII | 0.18 | — |
渔业资源评估模型采用动态产量模型(Dynamic Production Model)[19],参数估计基于以下6种假设:
(1)假设为复合种群,估计每一种群的K,q,r,B1997参数及种群间交换矩阵参数,该假设与模拟的种群动态一致。
(2)假设两个独立种群,分别估计两个种群的K,q,r,B1997参数。
(3)假设一个种群,合并捕捞产量,资源指数采用种群SI的CPUE,估计一个种群的K,q,r,B1997参数。
(4)假设一个种群,合并捕捞产量,资源指数采用种群SII的CPUE,估计一个种群的K,q,r,B1997参数。
(5)假设一个种群,合并捕捞产量,资源指数采用两亚种群的CPUE(采用相对CPUE,以去除捕捞系数的影响)按产量进行加权的平均值,估计一个种群的K,q,r,B1997参数。
F:假设一个种群,合并捕捞产量,资源指数采用合并的CPUE即式(4)计算值,估计一个种群的K,q,r,B1997参数。
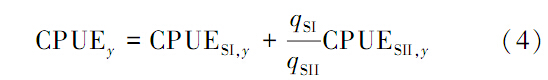
(1)管理目标
管理目标为获得最大可持续产量(MSY)。为此,根据估计的种群参数(Ki)定义管理目标生物量(B


若模型假设为单种群,则上式中i仅表示该种群。
(2)管理策略
根据管理目标定义的生物参考点(B
1)若B

当假设为两个独立种群或一个种群时,θi->j或θj->i为0。
2)若B

式中,CT
3)若B
此外,考虑采用更保守的F0.1作为捕捞控制目标[19, 21],以与MSY管理目标作对比分析。为此,将式(5)改为式(9),式(7)改为式(10),其它量的定义或计算方法不变。


在A、B假设条件下,I、II、III 3种管理策略,能直接计算两种群的捕捞量。但在C、D、E、F假设条件下,只能计算一个捕捞量,为此,本文采用两种方法将该捕捞量分配给两种群:
(1)利用式(2)计算CPUE,则两种群的捕捞量由式(11)、(12)计算,该方法假设管理者可以通过CPUE了解种群资源的空间分布状况。

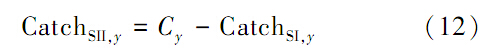
(2)平均分配捕捞量即CatchSI,y,CatchSII,y各为0.5C
由于有6种资源评估模型假设,C、D、E、F假设分别有2种捕捞量分配方法,因此共有10种评估、管理方案,分别标记为A、B、C1、C2、D1、D2、E1、E2、F1、F2,其中,字母分别表示6种假设,1与2分别对应方法(1)、(2)两种捕捞量分配方案。
1.3 结果评价指标根据管理策略计算的捕捞量及种群参数(表 1,表 2)即可推演种群动态过程,若种群生物量少于捕捞量,则捕捞量调整为此时生物量。假设管理期为10a。若10a后生物量小于或等于0,则该种群灭绝。本文将以下4个量作为评价资源评估及管理方案的指标:
(1)种群灭绝率,即计算2000次模拟当中, 两种群灭绝的比例;
(2)管理期平均捕捞量;
(3)管理期最后一年捕捞量,以考虑种群的恢复状况;
(4)10年管理期后,第1年2种群总生物量。
MSY可由式(7)计算,维持MSY时生物量(BMSY)可由式(5)计算,结果见表 1。本文95%置信区间采用统计量的2.5%与97.5%分位数区间表示。
2 结果模拟情况1—4,K与r均最大(表 1)。模拟情况1中,A、B、D1、D2、E1、E2、F1、F2方案均未出现种群灭绝,而对于C1与C2方案,两种群出现100%灭绝(表 3);A方案年均捕捞量为MSY(表 4),其10a后生物量为171×104t(表 5,平均值),大于BMSY(表 1);B方案出现过捕捞现象即年均捕捞量(74.3×104t)大于MSY,而10a后生物量(114×104t)小于BMSY;比较年均捕捞量与10a后总生物量,D1、E1、F1方案类似,均表现为开发不足,而D2、E2、F2方案分别与D1、E1、F1方案的捕捞量完全一致,但D2、E2、F2方案的10a后生物量均分别小于D1、E1、F1方案的10a后生物量,其中E2、F2方案的10a后资源量分别为143×104t与152×104t,小于BMSY(表 1,表 5),即尽管其年均捕捞量小于MSY,但仍出现过度捕捞现象(表 1,表 4,表 5)。模拟情况2与模拟情况1种群动态完全一致,仅CPUE数据随观测误差增大而质量变差。模拟情况2与模拟情况1的结果相比,A、B、E2、F2方案均使种群出现较大灭绝率,而在C1与C2方案下,其种群灭绝率有所降低,但在所有方案中仍较大,D1,D2、E1、F1方案下的种群基本不灭绝(表 3);从年均捕捞量与10a后生物量看,其2.5%与97.5%分位数区间变大,增加了结果的不确定性,方案差异显著性减弱(表 4,表 5)。模拟情况3、4分别与对应模拟情况1、2相比,其差异是种群空间交换率有所增大(表 1);模拟情况3、4下,A、C1、C2、D1、E1、E2、F1、F2方案的结果分别与模拟情况1、2类似,但对于B方案,种群出现了95%以上的灭绝率(表 3—表 5);与模拟情况2相比, D2方案在模拟情况4下出现较大种群灭绝率(表 3)。
|
方案
Schemes | 模拟情况 Scenarios | ||||||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | ||
| A | SI | 0.00 | 0.34 | 0.00 | 0.54 | 0.00 | 0.49 | 0.00 | 0.44 | 0.00 | 0.40 | 0.00 | 0.64 |
| SII | 0.00 | 0.07 | 0.00 | 0.26 | 0.00 | 0.32 | 0.00 | 0.22 | 0.00 | 0.09 | 0.00 | 0.42 | |
| B | SI | 0.00 | 0.53 | 1.00 | 0.95 | 1.00 | 0.76 | 1.00 | 0.96 | 0.00 | 0.60 | 1.00 | 0.99 |
| SII | 0.00 | 0.17 | 1.00 | 0.96 | 1.00 | 0.91 | 1.00 | 0.97 | 0.00 | 0.31 | 1.00 | 0.99 | |
| C1 | SI | 1.00 | 0.88 | 1.00 | 0.99 | 1.00 | 0.99 | 0.99 | 0.95 | 1.00 | 0.98 | 1.00 | 0.99 |
| SII | 1.00 | 0.89 | 1.00 | 0.99 | 1.00 | 0.99 | 0.99 | 0.95 | 1.00 | 0.98 | 1.00 | 0.99 | |
| C2 | SI | 1.00 | 0.80 | 1.00 | 1.00 | 1.00 | 0.95 | 0.99 | 0.95 | 1.00 | 0.91 | 1.00 | 0.99 |
| SII | 1.00 | 0.98 | 1.00 | 1.00 | 1.00 | 1.00 | 0.99 | 0.97 | 1.00 | 1.00 | 1.00 | 1.00 | |
| D1 | SI | 0.00 | 0.00 | 0.00 | 0.03 | 1.00 | 0.92 | 0.00 | 0.11 | 0.00 | 0.04 | 1.00 | 0.77 |
| SII | 0.00 | 0.00 | 0.00 | 0.04 | 1.00 | 0.92 | 0.00 | 0.11 | 0.00 | 0.05 | 1.00 | 0.77 | |
| D2 | SI | 0.00 | 0.00 | 0.00 | 0.05 | 0.00 | 0.38 | 0.00 | 0.09 | 0.00 | 0.02 | 1.00 | 0.80 |
| SII | 0.00 | 0.00 | 0.00 | 0.18 | 1.00 | 1.00 | 1.00 | 0.81 | 1.00 | 0.45 | 1.00 | 0.98 | |
| E1 | SI | 0.00 | 0.07 | 0.00 | 0.08 | 1.00 | 0.77 | 1.00 | 0.71 | 0.00 | 0.18 | 1.00 | 0.78 |
| SII | 0.00 | 0.06 | 0.00 | 0.09 | 1.00 | 0.76 | 1.00 | 0.71 | 0.00 | 0.18 | 1.00 | 0.78 | |
| E2 | SI | 0.00 | 0.03 | 0.00 | 0.12 | 0.00 | 0.53 | 1.00 | 0.70 | 0.00 | 0.10 | 1.00 | 0.79 |
| SII | 0.00 | 0.31 | 0.00 | 0.46 | 1.00 | 0.92 | 1.00 | 0.82 | 0.99 | 0.67 | 1.00 | 0.90 | |
| F1 | SI | 0.00 | 0.04 | 0.00 | 0.08 | 0.00 | 0.49 | 0.00 | 0.27 | 0.00 | 0.16 | 0.00 | 0.54 |
| SII | 0.00 | 0.04 | 0.00 | 0.08 | 0.00 | 0.49 | 0.00 | 0.27 | 0.00 | 0.16 | 0.00 | 0.55 | |
| F2 | SI | 0.00 | 0.02 | 0.00 | 0.10 | 0.00 | 0.20 | 0.00 | 0.25 | 0.00 | 0.11 | 0.00 | 0.56 |
| SII | 0.00 | 0.20 | 0.00 | 0.26 | 1.00 | 0.73 | 1.00 | 0.56 | 0.00 | 0.48 | 0.76 | 0.72 | |
模拟情况5—8较之于模拟情况1—4,其种群SII的K有所减少(表 1)。在模拟情况5—8下,A、C1与C2方案的种群灭绝率等结果与模拟情况1—4类似;不同点在于:与模拟情况1相比,在模拟情况5下,B、D1、E1方案的两种群及D2、E2、F2方案的种群SII出现了100%灭绝,随CPUE观测误差的变大即在模拟情况6下,采用B、D1、D2、E1、E2、F2方案的种群灭绝率有所降低,但仍维持在较高水平;与模拟情况3相比,在模拟情况7下,E1、E2方案的两种群、D2、F2方案的种群SII的灭绝率均为100%,尽管在模拟情况8下,灭绝率有所下降,但同样维持在较高水平;D1与F1方案,在CPUE观测误差较大时(模拟情况6、8)均有较大的种群灭绝率(表 3)。
模拟情况9—12较之于模拟情况1—4,其种群SII的r有所减少(表 1)。A、B、C1与C2方案下的种群灭绝率等结果与模拟情况1—4类似,不同点在于:模拟情况9,D2、E2方案的种群SII出现了99%以上的种群灭绝率;模拟情况11,D1、D2、E1与E2方案,两种群均出现100%的种群灭绝率,F2的种群灭绝率也达76%,在模拟情况12时,上述方案的种群灭绝率仍大于70%;F1在CPUE观测误差较大时(模拟情况10、12),种群灭绝率有较大提高(表 3)。
当采用F0.1作为捕捞控制目标时,各方案的种群灭绝率有不同程度的下降,其中D1与F1方案的种群灭绝率有大幅减少,且D1方案不存在100%种群灭绝率,但除A,D1,F1方案之外,其它方案仍存在100%种群灭绝率的情况(表 6)。
|
模拟情况
Scenarios | 统计量
Statistic | 方案Schemes | |||||||||
| A | B | C1 | C2 | D1 | D2 | E1 | E2 | F1 | F2 | ||
| 2.5%与97.5%为分位数;2.5% and 97.5% were quantile for each statistic; Mean was the average of the statistic | |||||||||||
| 1 | 平均 | 68.2 | 74.3 | 57.7 | 56.1 | 64.4 | 64.4 | 67.0 | 67.0 | 66.1 | 66.1 |
| 2.50% | 67.5 | 73.5 | 55.8 | 55.0 | 64.3 | 64.3 | 66.7 | 66.7 | 65.8 | 65.8 | |
| 97.50% | 68.9 | 75.1 | 59.3 | 57.8 | 64.5 | 64.5 | 67.3 | 67.3 | 66.4 | 66.4 | |
| 2 | 平均 | 63.3 | 65.0 | 52.4 | 51.2 | 65.3 | 65.3 | 68.5 | 67.1 | 67.6 | 66.7 |
| 2.50% | 31.7 | 44.0 | 30.0 | 29.9 | 62.2 | 62.2 | 62.7 | 62.7 | 62.5 | 62.4 | |
| 97.50% | 75.9 | 76.8 | 77.1 | 69.3 | 69.1 | 69.1 | 76.7 | 71.0 | 76.3 | 70.9 | |
| 3 | 平均 | 67.0 | 40.5 | 32.7 | 31.2 | 60.0 | 60.0 | 63.8 | 63.8 | 61.8 | 61.8 |
| 2.50% | 65.8 | 39.7 | 32.1 | 30.8 | 59.9 | 59.9 | 63.6 | 63.6 | 61.4 | 61.4 | |
| 97.50% | 68.2 | 41.2 | 33.1 | 31.5 | 60.1 | 60.1 | 64.1 | 64.1 | 62.1 | 62.1 | |
| 4 | 平均 | 53.2 | 41.3 | 32.1 | 31.2 | 62.6 | 61.3 | 64.5 | 61.5 | 61.8 | 60.2 |
| 2.50% | 20.5 | 24.8 | 22.9 | 23.7 | 58.6 | 55.1 | 50.5 | 45.4 | 38.0 | 36.3 | |
| 97.50% | 73.3 | 70.4 | 49.6 | 44.2 | 71.8 | 65.2 | 72.4 | 65.4 | 71.9 | 65.4 | |
| 5 | 平均 | 57.3 | 48.9 | 29.6 | 32.9 | 40.8 | 43.3 | 40.8 | 43.3 | 58.5 | 43.7 |
| 2.50% | 56.9 | 47.5 | 29.4 | 32.3 | 39.8 | 43.2 | 39.1 | 43.2 | 57.7 | 43.3 | |
| 97.50% | 57.7 | 50.7 | 30 | 33.4 | 42.1 | 43.4 | 42.6 | 43.5 | 59.3 | 44.6 | |
| 6 | 平均 | 49.9 | 45.4 | 29.5 | 32.0 | 41.0 | 41.3 | 38.9 | 38.3 | 46.3 | 43.7 |
| 2.50% | 32.2 | 32.1 | 21.9 | 22.8 | 28.5 | 31.4 | 21.5 | 22.4 | 26.8 | 29.3 | |
| 97.50% | 60.3 | 59.8 | 42.8 | 43.7 | 59.6 | 45.2 | 59.8 | 51.6 | 59.6 | 52.2 | |
| 7 | 平均 | 53.6 | 27.5 | 18.5 | 19.1 | 52.5 | 46.4 | 27.0 | 26.5 | 51.8 | 46.0 |
| 2.50% | 53.3 | 27.2 | 18.2 | 18.8 | 52.3 | 46.2 | 25.0 | 24.1 | 51.4 | 45.3 | |
| 97.50% | 53.9 | 28.0 | 18.5 | 19.1 | 52.6 | 46.5 | 28.8 | 27.9 | 52.1 | 47.8 | |
| 8 | 平均 | 46.1 | 28.2 | 19.5 | 19.6 | 51.8 | 44.9 | 31.4 | 29.8 | 45.2 | 41.1 |
| 2.50% | 24.6 | 20.5 | 14.1 | 14.8 | 39.2 | 34.5 | 16.8 | 16.4 | 23.3 | 23.4 | |
| 97.50% | 56.7 | 46.4 | 47.3 | 45.2 | 55.3 | 51.2 | 54.2 | 49.8 | 54.7 | 50.3 | |
| 9 | 平均 | 64.8 | 71.0 | 43.9 | 45.0 | 64.6 | 63.6 | 63.5 | 62.8 | 61.3 | 61.3 |
| 2.50% | 64.0 | 70.5 | 43.0 | 44.2 | 64.3 | 63.4 | 63.1 | 62.4 | 60.8 | 60.8 | |
| 97.50% | 65.5 | 71.6 | 45.1 | 45.6 | 64.9 | 63.7 | 63.8 | 63.0 | 61.7 | 61.7 | |
| 10 | 平均 | 59.9 | 60.7 | 41.7 | 41.9 | 63.7 | 62.0 | 62.3 | 58.5 | 60.0 | 58.0 |
| 2.50% | 34.5 | 37.7 | 25.2 | 25.3 | 59.2 | 57.8 | 36.3 | 37.9 | 31.1 | 30.7 | |
| 97.50% | 70.7 | 71.5 | 68.1 | 57.2 | 70.2 | 64.1 | 70.8 | 63.0 | 69.8 | 63.0 | |
| 11 | 平均 | 60.7 | 34.1 | 24.5 | 24.4 | 51.8 | 40.6 | 40.0 | 35.5 | 56.8 | 51.7 |
| 2.50% | 60.2 | 33.6 | 24.1 | 24 | 50.6 | 40.4 | 35.1 | 32.4 | 52.9 | 46.7 | |
| 97.50% | 61.1 | 34.5 | 24.8 | 24.7 | 53.4 | 40.8 | 45.6 | 38.6 | 59.4 | 55.7 | |
| 12 | 平均 | 48.3 | 32.2 | 23.4 | 23.3 | 49.2 | 40.9 | 36.8 | 33.8 | 45.2 | 41.1 |
| 2.50% | 25.6 | 21.3 | 17.8 | 17.8 | 32.8 | 31.5 | 19.1 | 19.4 | 22.6 | 23.1 | |
| 97.50% | 64.1 | 46.2 | 31.6 | 30.9 | 61.8 | 53.8 | 60.8 | 55.6 | 61.2 | 55.1 | |
|
模拟情况
Scenarios | 统计量
Statistic | 方案Schemes | |||||||||
| A | B | C1 | C2 | D1 | D2 | E1 | E2 | F1 | F2 | ||
| 2.5%与97.5%为分位数;2.5% and 97.5% were quantile for each statistic; Mean was the average of the statistic | |||||||||||
| 1 | 平均 | 17.1 | 11.4 | 0.0 | 0.0 | 18.2 | 16.4 | 16.9 | 14.3 | 17.5 | 15.2 |
| 2.50% | 16.6 | 10.2 | 0.0 | 0.0 | 18.0 | 16.3 | 16.7 | 13.9 | 17.2 | 14.9 | |
| 97.50% | 17.5 | 12.3 | 0.0 | 0.0 | 18.3 | 16.5 | 17.2 | 14.6 | 17.7 | 15.5 | |
| 2 | 平均 | 11.7 | 7.3 | 0.9 | 0.9 | 17.4 | 15.5 | 13.9 | 11.5 | 15.0 | 12.7 |
| 2.50% | 0.9 | 0.0 | 0.0 | 0.0 | 14.9 | 11.5 | 0.0 | 0.0 | 0.0 | 2.2 | |
| 97.50% | 18.9 | 17.9 | 12.9 | 7.8 | 18.9 | 17.6 | 19.0 | 17.5 | 19.1 | 17.7 | |
| 3 | 平均 | 16.4 | 0.0 | 0.0 | 0.0 | 18.1 | 15.2 | 16.5 | 10.6 | 17.7 | 13.7 |
| 2.50% | 15.5 | 0.0 | 0.0 | 0.0 | 17.9 | 15.1 | 16.2 | 10.1 | 17.5 | 13.3 | |
| 97.50% | 17.1 | 0.0 | 0.0 | 0.0 | 18.3 | 15.3 | 16.7 | 11.1 | 17.9 | 14.1 | |
| 4 | 平均 | 8.4 | 0.5 | 0.1 | 0.0 | 15.6 | 11.6 | 13.1 | 7.7 | 14.6 | 10.4 |
| 2.50% | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| 97.50% | 18.8 | 9.2 | 0.0 | 0.0 | 18.7 | 16.1 | 18.1 | 15.0 | 18.7 | 16.2 | |
| 5 | 平均 | 13.7 | 0.0 | 0.0 | 0.0 | 0.0 | 2.7 | 0.0 | 2.7 | 10.0 | 7.5 |
| 2.50% | 13.2 | 0.0 | 0.0 | 0.0 | 0.0 | 2.2 | 0.0 | 1.6 | 7.4 | 7.2 | |
| 97.50% | 14.1 | 0.0 | 0.0 | 0.0 | 0.0 | 3.3 | 0.0 | 3.5 | 12.1 | 7.8 | |
| 6 | 平均 | 7.1 | 1.4 | 0.1 | 0.2 | 0.7 | 2.6 | 2.9 | 3.3 | 7.1 | 7.0 |
| 2.50% | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| 97.50% | 16.2 | 12.2 | 0.0 | 3.1 | 11.6 | 7.9 | 16.6 | 14.7 | 17.6 | 16.3 | |
| 7 | 平均 | 13.7 | 0.0 | 0.1 | 0.1 | 13.5 | 5.4 | 0.0 | 0.0 | 14.0 | 5.5 |
| 2.50% | 13.4 | 0.0 | 0.0 | 0.0 | 13.2 | 5.2 | 0.0 | 0.0 | 13.5 | 5.0 | |
| 97.50% | 14.1 | 0.0 | 0.0 | 0.0 | 13.7 | 5.6 | 0.0 | 0.0 | 14.5 | 6.1 | |
| 8 | 平均 | 7.8 | 0.5 | 0.7 | 0.6 | 10.5 | 5.1 | 4.0 | 2.9 | 10.4 | 7.4 |
| 2.50% | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| 97.50% | 16.7 | 11.6 | 16.8 | 13.6 | 15.9 | 12.9 | 17.2 | 15.7 | 18.0 | 16.8 | |
| 9 | 平均 | 16.1 | 6.4 | 0.0 | 0.0 | 13.7 | 8.3 | 15.7 | 8.9 | 17.3 | 12.6 |
| 2.50% | 15.4 | 4.8 | 0.0 | 0.0 | 13.3 | 8.1 | 15.3 | 8.7 | 16.9 | 11.8 | |
| 97.50% | 16.6 | 7.8 | 0.0 | 0.0 | 14.1 | 8.5 | 16.1 | 9.1 | 17.6 | 13.5 | |
| 10 | 平均 | 10.3 | 5.0 | 0.2 | 0.3 | 13.1 | 9.5 | 11.1 | 8.0 | 12.7 | 9.5 |
| 2.50% | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.4 | 0.0 | 0.0 | 0.0 | 0.0 | |
| 97.50% | 18.1 | 16.4 | 0.1 | 4.5 | 17.8 | 14.6 | 18.0 | 15.5 | 18.4 | 16.3 | |
| 11 | 平均 | 15.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 15.1 | 6.4 |
| 2.50% | 14.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 10.8 | 2.7 | |
| 97.50% | 15.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 18.0 | 14.2 | |
| 12 | 平均 | 5.9 | 0.1 | 0.1 | 0.1 | 2.1 | 0.7 | 2.8 | 1.6 | 6.5 | 4.5 |
| 2.50% | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| 97.50% | 17.8 | 0.0 | 0.0 | 0.0 | 15.3 | 6.7 | 17.5 | 14.2 | 18.7 | 16.6 | |
|
方案
Schemes | 模拟情况 Scenarios | ||||||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | ||
| A | SI | 0.00 | 0.24 | 0.00 | 0.41 | 0.00 | 0.19 | 0.00 | 0.21 | 0.00 | 0.21 | 0.00 | 0.39 |
| SII | 0.00 | 0.10 | 0.00 | 0.30 | 0.00 | 0.09 | 0.00 | 0.19 | 0.00 | 0.03 | 0.00 | 0.28 | |
| B | SI | 0.00 | 0.32 | 1.00 | 0.83 | 0.00 | 0.23 | 1.00 | 0.85 | 0.00 | 0.30 | 1.00 | 0.92 |
| SII | 0.00 | 0.00 | 1.00 | 0.81 | 0.00 | 0.21 | 1.00 | 0.89 | 0.00 | 0.00 | 1.00 | 0.95 | |
| C1 | SI | 0.76 | 0.67 | 1.00 | 0.98 | 1.00 | 0.95 | 0.99 | 0.95 | 1.00 | 0.84 | 1.00 | 0.99 |
| SII | 0.79 | 0.66 | 1.00 | 0.98 | 1.00 | 0.95 | 0.99 | 0.95 | 1.00 | 0.85 | 1.00 | 0.99 | |
| C2 | SI | 0.00 | 0.56 | 1.00 | 0.99 | 0.98 | 0.64 | 0.99 | 0.94 | 1.00 | 0.68 | 1.00 | 0.99 |
| SII | 1.00 | 0.85 | 1.00 | 0.99 | 1.00 | 1.00 | 0.99 | 0.96 | 1.00 | 0.99 | 1.00 | 0.99 | |
| D1 | SI | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.40 | 0.00 | 0.00 | 0.00 | 0.01 | 0.00 | 0.18 |
| SII | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.40 | 0.00 | 0.00 | 0.00 | 0.01 | 0.00 | 0.18 | |
| D2 | SI | 0.00 | 0.00 | 0.00 | 0.01 | 0.00 | 0.05 | 0.00 | 0.00 | 0.00 | 0.01 | 0.00 | 0.20 |
| SII | 0.00 | 0.00 | 0.00 | 0.04 | 1.00 | 0.97 | 0.00 | 0.21 | 0.00 | 0.08 | 1.00 | 0.86 | |
| E1 | SI | 0.00 | 0.01 | 0.00 | 0.03 | 0.00 | 0.54 | 1.00 | 0.61 | 0.00 | 0.07 | 0.58 | 0.63 |
| SII | 0.00 | 0.01 | 0.00 | 0.03 | 0.00 | 0.54 | 1.00 | 0.61 | 0.00 | 0.07 | 0.59 | 0.64 | |
| E2 | SI | 0.00 | 0.00 | 0.00 | 0.03 | 0.00 | 0.28 | 1.00 | 0.58 | 0.00 | 0.05 | 0.73 | 0.64 |
| SII | 0.00 | 0.05 | 0.00 | 0.10 | 1.00 | 0.85 | 1.00 | 0.76 | 0.00 | 0.22 | 1.00 | 0.82 | |
| F1 | SI | 0.00 | 0.00 | 0.00 | 0.05 | 0.00 | 0.21 | 0.00 | 0.10 | 0.00 | 0.09 | 0.00 | 0.35 |
| SII | 0.00 | 0.01 | 0.00 | 0.05 | 0.00 | 0.22 | 0.00 | 0.10 | 0.00 | 0.09 | 0.00 | 0.35 | |
| F2 | SI | 0.00 | 0.00 | 0.00 | 0.05 | 0.00 | 0.06 | 0.00 | 0.07 | 0.00 | 0.07 | 0.00 | 0.36 |
| SII | 0.00 | 0.03 | 0.00 | 0.09 | 1.00 | 0.62 | 0.00 | 0.36 | 0.00 | 0.18 | 0.00 | 0.60 | |
动态产量模型所要求的数据简单、比较适合对短寿命种类的资源进行评估[14]。目前,东、黄海日本鲐主要由0至2岁群体组成[22],尽管日本收集了捕捞年龄数据,但我国灯光围网渔业缺乏相应数据,因此目前采用动态产量模型对该资源进行评估较为合适。由于日本鲐环境承载容量(K)[22]、内禀增长率(r)[22]、空间交换矩阵等参数信息仍不足,我国(大陆)鲐鱼产量数据并没有区分日本鲐与澳洲鲐(Scomber australasicus),并且中、日、韩等国家或地区的日本鲐捕捞海域、种群归属等信息也基本缺失。因此本文模拟假设与实际东、黄海日本鲐的种群动态不同,但这并不妨碍本文展示种群的空间结构对渔业资源评估与管理的影响及管理策略评价的重要性(MSE)[20]。
评估管理方案A与模拟的种群动态一致。方案B考虑了种群的空间分布,假设了两个种群,但忽略了种群间的空间交换。C1、D1、E1、F1、C2、D2、E2、F2方案均将日本鲐资源假设为一个种群,C、D、E与F的差异仅在于选择了不同的CPUE,C、D、E的CPUE均反映部分种群动态信息,但F方案的CPUE能正确反映整个种群动态变化。对于C1、D1、E1与F1方案,管理者需要了解种群的空间结构并按比例分配捕捞量,而对于C2、D2、E2与F2方案,管理者忽略种群的空间结构而均匀分配捕捞量。
当CPUE观测误差较小情况下,A方案能获得MSY(表 7),10a后生物量大于或接近BMSY(表 1,表 5)。但随CPUE观测误差增大,A方案种群灭绝率增大,管理效果随之退化(表 3—表 5)。
B、C1、C2方案均存在过度开发,具有较大的种群灭绝率。D1、D2、E1与E2方案的种群灭绝率与K、r、种群空间交换率等参数有关。D1与D2、E1与E2参数估计方法完全一致,但对比种群灭绝率、年均捕捞量及10a后生物量,D1、E1方案总体上分别优于D2、E2方案。如模拟情况1下,D1与D2方案的年均捕捞量完全一致,但10a后的生物量,D1方案的值大于D2(表 4,表 5)。
F1方案与A方案类似,均不存在100%的种群灭绝率,而其它方案则有可能导致种群100%灭绝(与种群参数有关),如D1方案在模拟情况5、11时,均导致种群100%灭绝。虽然F1方案均呈现开发不足、不能获得MSY(表 7),但其种群灭绝率较低且不受种群参数影响,因此仍可作为A方案的替代方案,即若缺少种群信息,但能获得反映整个资源变化趋势的资源指数并使其精度得到保证,在此条件下,假设单一种群进行资源评估仍可接受,但必须将捕捞量按种群的空间结构进行分配,因为F2方案同样有种群100%灭绝(模拟情况5、7,表 3)。由于不同渔业其捕捞位置及捕捞种群存在差异,其CPUE数据可能仅反映渔业资源的部分信息,在渔业资源评估中,常采用不同的平均方法以期获得能反映资源总体变化趋势的资源指数[23]如式(4)和假设E,但该方法仍存在较大随意性。
|
模拟情况
Scenarios | 统计量
Statistic | 方案Schemes | |||||||||
| A | B | C1 | C2 | D1 | D2 | E1 | E2 | F1 | F2 | ||
| 2.5%与97.5%为分位数;2.5% and 97.5% were quantile for each statistic; Mean was the average of the statistic | |||||||||||
| 1 | 平均 | 68.2 | 74.3 | 0.0 | 0.0 | 65.0 | 65.0 | 67.0 | 67.0 | 66.1 | 66.1 |
| 2.50% | 67.5 | 73.6 | 0.0 | 0.0 | 64.9 | 64.9 | 66.7 | 66.7 | 65.8 | 65.8 | |
| 97.50% | 68.9 | 75.1 | 0.0 | 0.0 | 65.1 | 65.1 | 67.3 | 67.3 | 66.4 | 66.4 | |
| 2 | 平均 | 52.1 | 47.3 | 11.0 | 10.1 | 65.8 | 65.8 | 65.2 | 59.2 | 65.6 | 61.6 |
| 2.50% | 1.9 | 0.0 | 0.0 | 0.0 | 63.5 | 63.5 | 0.0 | 36.4 | 9.3 | 36.5 | |
| 97.50% | 75.7 | 76.5 | 76.6 | 63.6 | 69.6 | 69.6 | 75.7 | 70.8 | 76.0 | 70.7 | |
| 3 | 平均 | 68.2 | 0.0 | 0.1 | 0.0 | 62.0 | 62.0 | 63.8 | 63.8 | 62.0 | 62.0 |
| 2.50% | 67.0 | 0.0 | 0.0 | 0.0 | 61.8 | 61.8 | 63.6 | 63.6 | 61.8 | 61.8 | |
| 97.50% | 69.5 | 0.0 | 0.0 | 0.0 | 62.2 | 62.2 | 64.1 | 64.1 | 62.1 | 62.1 | |
| 4 | 平均 | 32.9 | 3.8 | 0.6 | 0.3 | 62.5 | 57.9 | 60.3 | 48.8 | 59.5 | 54.2 |
| 2.50% | 0.0 | 0.0 | 0.0 | 0.0 | 34.4 | 3.2 | 0.0 | 0.0 | 0.0 | 0.0 | |
| 97.50% | 74.5 | 68.9 | 0.0 | 0.0 | 71.9 | 68.2 | 71.6 | 65.3 | 71.8 | 65.9 | |
| 5 | 平均 | 60.5 | 0.1 | 0.0 | 0.0 | 0.0 | 32.5 | 0.0 | 32.5 | 59.5 | 29.8 |
| 2.50% | 60.3 | 0.0 | 0.0 | 0.0 | 0.0 | 32.3 | 0.0 | 32.2 | 59.1 | 29.5 | |
| 97.50% | 60.7 | 0.0 | 0.0 | 0.0 | 0.0 | 32.7 | 0.0 | 32.9 | 60.0 | 30.0 | |
| 6 | 平均 | 36.0 | 13.1 | 0.9 | 2.2 | 5.5 | 21.7 | 14.4 | 18.2 | 30.7 | 33 |
| 2.50% | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| 97.50% | 74.5 | 61.3 | 0.0 | 32.9 | 60.3 | 33.2 | 61.4 | 60.8 | 64.0 | 63.7 | |
| 7 | 平均 | 60.5 | 0.0 | 0.3 | 0.3 | 57.4 | 28.7 | 0.0 | 0.0 | 56.4 | 28.2 |
| 2.50% | 60.0 | 0.0 | 0.0 | 0.0 | 57.4 | 28.7 | 0.0 | 0.0 | 56.3 | 28.1 | |
| 97.50% | 60.9 | 0.0 | 0.0 | 0.0 | 57.5 | 28.7 | 0.0 | 0.0 | 56.6 | 28.3 | |
| 8 | 平均 | 37.5 | 2.5 | 2.7 | 2.4 | 52.6 | 34.5 | 16.9 | 14.4 | 42.1 | 34.9 |
| 2.50% | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| 97.50% | 71.5 | 55.0 | 55.1 | 54.2 | 58.9 | 57.6 | 60.9 | 60.9 | 60.0 | 60.0 | |
| 9 | 平均 | 65.1 | 70.9 | 0.0 | 0.0 | 68.6 | 58.6 | 63.5 | 56.4 | 61.6 | 61.6 |
| 2.50% | 64.6 | 70.5 | 0.0 | 0.0 | 68.2 | 54.8 | 63.1 | 49.6 | 61.5 | 61.5 | |
| 97.50% | 65.7 | 71.6 | 0.0 | 0.0 | 69.1 | 62.3 | 63.8 | 62.3 | 61.7 | 61.7 | |
| 10 | 平均 | 48.9 | 38.7 | 2.1 | 4.4 | 65.7 | 56.0 | 54.4 | 42.4 | 54.4 | 47.2 |
| 2.50% | 0.0 | 0.0 | 0.0 | 0.0 | 15.5 | 33.4 | 0.0 | 0.0 | 0.0 | 0.0 | |
| 97.50% | 73.3 | 71.4 | 47.8 | 37.8 | 79.7 | 69.9 | 70.7 | 63.6 | 70.9 | 67.4 | |
| 11 | 平均 | 64.9 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 58.6 | 38.6 |
| 2.50% | 64.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 57.7 | 29.2 | |
| 97.50% | 65.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 59.7 | 58.1 | |
| 12 | 平均 | 27.4 | 0.5 | 0.3 | 0.2 | 15.9 | 9.0 | 13.8 | 10.4 | 28.1 | 23.1 |
| 2.50% | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| 97.50% | 81.1 | 0.0 | 0.0 | 0.0 | 61.8 | 47.3 | 63.5 | 63.1 | 67.7 | 67.4 | |
在东、黄海日本鲐资源评估中,日本鲐均被假设为单一种群。陈卫忠等[24]、张洪亮等[16]根据局部海域(东海或浙江近海)捕捞产量及CPUE数据对相应海域日本鲐进行资源评估,其假设类似B,李纲等[17]虽然考虑了日本等国家或地区的产量数据,但其CPUE数据来自中国(大陆)大型灯光围网渔业数据,其假设类似C或D,日本学者使用日本与韩国的捕捞产量及相应CPUE数据[18]进行资源评估,其假设也类似B。尽管假设E或F并未出现在东、黄海日本鲐资源评估中,但在其它资源评估中常被使用[23]。从东、黄海日本鲐管理措施研究来看,目前仅提出总许可渔获量(Total Allowable Catch,TAC),并未考虑基于种群空间结构的TAC管理方案[17, 18]。
目前,对东、黄海日本鲐进行种群鉴别及按标准收集相关数据仍很困难,A方案没有实现可能,况且CPUE等数据的精确度难以保证,A方案也难以保证为最佳方案。若按现有的资源评估方法及以获得最大可持续产量作为管理目标,如模拟结果所示(B、C1、C2、D1、D2方案),东、黄海日本鲐可能存在较大的种群灭绝概率。若将F0.1作为捕捞控制目标,各方案种群灭绝率有所改善,特别是 D1,F1方案的种群灭绝率有较大减少,但D2、F2方案仍存在种群100%灭绝的情况(表 6)。因此,基于目前日本鲐生态学研究及数据现状,要使日本鲐资源可持续利用,必须设定更保守的捕捞量和采用基于种群空间结构的TAC管理方案。基于种群空间结构的TAC渔业管理已引起渔业学者和渔业资源管理者的重视,如在大西洋鲱鱼的TAC管理中,捕捞量的控制已考虑了种群的空间分布状况[25]。
由于渔业资源评估模型均存在一定假设,如假设环境承载容量、内禀增长率不受环境等因素影响,忽略种群的空间结构等[26]。收集的渔业数据也存在不确定性如捕捞数据存在瞒报、丢弃等现象[26],CPUE数据则可能存在较大误差。如上所述,当CPUE误差较小时,模型假设正确与否对管理效果有重要影响,正确的模型假设能得到最优的管理效果如A方案。但随CPUE误差增大,各方案管理效果的差异逐步缩小,不确定性增大(表 3—表 5)。因此根据估计参数设计生物参考点,进行渔业管理,可能会因模型假设,数据收集精度等问题导致风险。如本文,根据模型假设及估计的参数而设定的捕捞量均能满足相应假设下资源可持续利用要求,但若在真实的种群动态下,则可能会导致种群的灭绝。本文仅对几种假设情况进行了分析,所得结论不一定具有一般性,在实际渔业管理中,应结合种群生态、数据、模型假设及参数估计方法等方面的不确定性对管理控制规则进行系统的管理策略评价[20]。
| [1] | Hilborn R, Walters C. Quantitative Fisheries Stock Assessment: Choice, Dynamics, and Uncertainty. New York: Chapman and Hall, 1992:67-69. |
| [2] | Gotelli N. A Primer of Ecology. 3rd ed. Massachusetts, Sunderland: Sinauer Associates, Incorporated, 2001: 236. |
| [3] | Kerr L A, Cadrin S X, Secor D H. The role of spatial dynamics in the stability, resilience, and productivity of an estuarine fish population. Ecological Applications, 2010, 20:497-507. |
| [4] | Wu J G. What is metapopulation, really? Acta Phytoecologica Sinica, 2000, 24(1):123-126. |
| [5] | Kritzer J P, Sale P F. Metapopulation ecology in the sea: from Levins' model to marine ecology and fisheries science. Fish and Fisheries, 2004, 5(2): 131-140. |
| [6] | Stephenson R L, Melvin G D, Power M J. Population integrity and connectivity in Northwest Atlantic herring: a review of assumptions and evidence. ICES Journal of Marine Science, 2009, 66: 1733-1739. |
| [7] | Zhang Q H, Chen J H, Xu H X, Shen X Q, Yu G P, Zheng Y J. The fishery resources in East China Sea and it's sustainable use. Shanghai: Fudan Press, 2007:213-215. |
| [8] | Zhu D S, Wang W X, Zhang G X. Studies on the fisheries biology of mackerel (Pneumatophorus japonicus Houttuyn) in the yellow sea Ⅰ on the migratory and distributional patterns of mackerel in the Yellow Sea and Bohai Sea. Marine Fisheries Research, 1982, 4:17-31. |
| [9] | Chen Q, Wang Y J, Li P J, Xiang F C, Liu C Y. Migration and distribution of chub mackerel Scomber japonicus in the Yellow Sea and Bohai Sea. Fisheries Science, 1983, 1:6-13. |
| [10] | Shao F, Chen X J, Li G. Morphological variations and discriminant analysis of Scomber japonicus and Scomber australasicus in the Yellow Sea and East China Sea. Journal of shanghai fisheries university, 2008, 17(2):204-209. |
| [11] | Li Y S. Simulation study of ecological dynamics of early life history of Chub mackerel (Scomber japonicus) in the East China Sea [D]. Shanghai: Shanghai Ocean University, 2012. |
| [12] | Guan W J, Chen X J, Li G. Influence of sea surface temperature and La Nina event on temporal and spatial fluctuation of chub mackerel (Scomber japonicus) stock in the East China Sea. Journal of shanghai ocean university, 2011, 20(1):102-107. |
| [13] | Cadrin S X, Secor D H. Accounting for spatial population structure in stock assessment: past, present and future//Beamish R J, Rothschild B J eds. The Future of Fishery Science in North America, Fish & Fisheries Series, Volume 31. Netherlands: Springer, 2009:405-426. |
| [14] | Ying Y, Chen Y, Lin L, Gao T. Risks of ignoring fish population spatial structure in fisheries management. Canadian Journal of Fisheries and Aquatic Sciences, 2011, 68(12): 2101-2120. |
| [15] | Stephenson R L. Stock complexity in fisheries management: a perspective of emerging issues related to population sub-units. Fisheries Research, 1999, 43: 247-249. |
| [16] | Zhang H L, Zhou Y D, Yao G Z. Analysis on the utilization of resources in small scale traditional light seining fisheries of Zhejiang Province. Marine Fisheries, 2007, 29(2): 174-178. |
| [17] | Li G, Chen X J, Guan W J. Stock assessment and risk analysis of management strategies for Scomber japonicus in the East China Sea and Yellow Sea using a Bayesian approach. Journal of Fisheries of China, 2010, 34(5):740-750. |
| [18] | Yukami R, Yoda M, Ohshimo S, Tanaka H, Kurota H. Stock assessment of Chub mackerel Scomber japonicus Tsushima Warm Current Stock in 2011.[2012-10-28]. |
| [19] | Quinn T J, Deriso R B. Quantitative Fish Dynamic. New York: Oxford University Press, 1999:398-401. |
| [20] | Punt A E, Ralston S. A management strategy evaluation of rebuilding revision rules for overfished rockfish stocks//Heifetz J, DiCosimo J, Gharrett A J, Love M S, O'Connell V M, Stanley R D, eds. Biology, Assessment, and Management of North Pacific Rockfishes. Alaska Sea Grant College Program, AK-SG-07-01, 2007: 329-351. |
| [21] | Chen Y, Montgomery S S. Modeling the dynamics of eastern rock lobster, Jasus verreauxi, stock in New South Wales, Australia. Fisheries Bulletin, 1999, 97: 25-38. |
| [22] | Li G. Stock Assessment and risk analysis of Chub mackerel (Scomber japonicus) in the East China Sea and the Yellow Sea [D]. Shanghai: Shanghai Fisheries University, 2008. |
| [23] | International Commission for the Conservation of Atlantic Tunas. Report of the Standing Committee on Research and Statistics (SCRS). Madrid: ICCAT, 2012. [2012-10-28]. |
| [24] | Chen W Z, Hu F, Yan L P. Stock assessment of scomber japoniscus from the east china sea with virtual population analysis. Journal of Fisheries of China, 1998, 22(4):334-339. |
| [25] | New England Fishery Management Council. Proposed Atlantic herring specifications for the 2010—2012 fishing years (January 1, 2010-December 31, 2012) including the environmental assessment (EA), regulatory impact review (RIR), and initial regulatory flexibility analysis (IRFA). Newburyport, Massachusetts: NEFMC, 2010. [2012-10-28]. |
| [26] | Guan W J, Gao F, Lei L, Chen X J. Respective problem in fishery stock assessment. Journal of Shanghai Ocean University, 2012, 21(5):841-847. |
| [4] | 邬建国. Metapopulation(复合种群)究竟研究是什么?. 植物生态学报, 2000,24(1):123-126. |
| [7] | 张秋华, 程家华, 徐汉祥, 沈新强, 俞国平, 郑元甲. 东海区渔业资源及可持续利用. 上海: 复旦大学出版社, 2007:213-215. |
| [8] | 朱德山, 王为祥, 张国祥. 黄海鲐鱼(Pneumatophorus japonicus Houtuyn)渔业生物学研究 I 黄、渤海鲐鱼洄游分布研究. 海洋水产研究, 1982,4: 17-31. |
| [9] | 陈俅, 王有君, 李培君, 项福椿, 刘春洋. 黄渤海区日本鲐的洄游与分布.水产科学学, 1983, 1: 6-13. |
| [10] | 邵锋, 陈新军, 李纲, 钱卫国. 东黄海鲐鱼形态差异分析.上海水产大学学报, 2008, 17 (2): 204-209. |
| [11] | 李曰嵩. 东海鲐鱼(Scomber japonica)早期生活史过程的生态动力学模拟研究[D]. 上海:上海海洋大学, 2012. |
| [12] | 官文江, 陈新军, 李纲. 海表水温和拉尼娜事件对东海鲐鱼资源及时空变动的影响.上海海洋大学学报, 2011, 20(1):102-107. |
| [16] | 张洪亮, 周永东, 姚光展. 浙江群众传统灯光围网渔业利用资源状况分析.海洋渔业, 2007, 29(2): 174-178. |
| [17] | 李纲, 陈新军, 官文江. 基于贝叶斯方法的东黄海鲐资源评估及管理策略风险分析.水产学报, 2010, 34(5):740-750. |
| [18] | 由上龍嗣,依田真里, 大下誠二, 田中寛繁, 黒田啓行. 平成23年マサバ対馬暖流系群の資源評価. [2012-10-28]. |
| [24] | 陈卫忠, 胡芬, 严利平. 用实际种群分析法评估东海鲐鱼现存资源量. 水产学报, 1998, 22(4):334-339. |
| [26] | 官文江, 高峰,雷林, 陈新军. 渔业资源评估中的回顾性问题.上海海洋大学学报, 2012, 21(5):841-847. |