 2014, Vol. 34
2014, Vol. 34文章信息
- 范夫静, 黄国勤, 宋同清, 曾馥平, 彭晚霞, 杜虎, 文丽, 何铁光
- FAN Fujing, HUANG Guoqin, SONG Tongqing, ZENG Fuping, PENG Wanxia, DU Hu, WEN Li, HE Tieguang
- 西南峡谷型喀斯特坡地土壤微生物量C、N、P空间变异特征
- Spatial heterogeneity of soil microbial biomass carbon, nitrogen, and phosphorus in sloping field in a groge Karst region, Southwest China
- 生态学报, 2014, 34(12): 3293-3301
- Acta Ecologica Sinica, 2014, 34(12): 3293-3301
- http://dx.doi.org/10.5846/stxb201310302612
-
文章历史
- 收稿日期:2013-10-30
- 修订日期:2014-4-24






2. 中国科学院亚热带农业生态研究所亚热带农业生态过程重点实验室, 长沙 410125;
3. 中国科学院环江喀斯特生态系统观测研究站, 环江 547100;
4. 广西农业科学研究院农业资源与环境研究所, 南宁 530004
2. Institute of Subtropical Agriculture, Chinese Academy of Sciences, Changsha 410125, China;
3. Karst Station for Ecosystem in Huanjiang, Institute of Subtropical Agriculture, Chinese Academy of Sciences, Huanjiang 547100, China;
4. Guangxi Agricultural Science Research Institute of Agricultural Resources And Environment Research Institute, Nanning 530004, China
土壤微生物是维持土壤品质的重要组成部分,几乎参与土壤中一切生物和生物化学反应,对土壤中的动植物残体和土壤有机质及其有害物质的分解、元素的地球生物化学循环和土壤结构的形成过程起着重要的调节作用。土壤微生物对养分循环与平衡、土壤理化性质的改善也起着重要调控作用[1, 2]。它对环境变化敏感,其呼吸强度是衡量土壤微生物总的活性指标[3, 4],而微生物量的任何变化,都会影响土壤养分循环和有效性[5, 6],能够较早地指示生态系统功能的变化。国内外关于土壤微生物量的研究主要集中在垂直变异、季节动态及影响因子等方面,获得了土壤微生物量上层一般高于下层、随着土壤深度的增加呈下降趋势、同一生态系统中不同植被土壤微生物量差异显著[7]等重要认识。土壤微生物作为退化生态系统恢复的“先锋者”,研究其空间格局对喀斯特坡地生态系统的恢复及重建具有重要的意义。
长期以来,不同学者提出了许多解释大型生物(动物和植物)空间分布格局形成和维持机制的假说和理论,推动了生物地理学的发展,但微生物生物地理学的研究十分薄弱,甚至对微生物是否存在一定的地理分布格局都存在广泛争议[8, 9]。喀斯特独特的地质生态环境决定了喀斯特土壤微生物的特异性,何寻阳等[10]、李新爱等[11]初步研究了喀斯特峰丛洼地的土壤微生物特征,但关于土壤微生物空间分布的报道甚少[12]。因此,本文选取西南峡谷型喀斯特坡地为研究对象,采用网格法采样,用经典统计学和地统计学方法分析土壤微生物生物量碳(MBC)、氮(MBN)、磷(MBP)的空间变异特征,揭示影响其空间格局的主要因子,为指导喀斯特退化生态系统植被的迅速恢复和生态重建、推动微生物生物地理学发展提供科学理论支持和参考。
1 研究方法 1.1 研究区概况研究区地处贵州省晴隆县孟寨河小流域,位于25°33′—26°11′N、105°01′—105°25′E之间,最高海拔2025 m,因受北盘江及其支流的强烈切割,切深长达500—700 m,属深切割岩溶侵蚀高原峡谷区。亚热带湿润季风气候特点较为明显,气候温和湿润,年平均气温14.0—15.9 ℃,年极端高温为33.4 ℃,最低气温为-6.2℃,日照时数1453 h,无霜期280 d左右。年降水量1 500—1 650 mm,集中在6—9月份,年蒸发量1800 mm,空气相对湿度在50%以下。森林覆盖率为25.1%,林草植被覆盖度为27.8%,森林植被以阔叶林为主,针叶林次之。原生植被基本上被破坏,现为次生林。该地区地质结构复杂,地形起伏大,具有山高、谷深、坡陡的特点,属二迭纪岩层风化而成的石灰土。
1.2 样品采集与分析 1.2.1 样品采集根据典型性、代表性原则,经反复勘查,在研究区内选取面积为300 m × 200 m的典型峡谷型喀斯特坡地,该坡地2008年退耕种草放羊,由于存在地表地下双层空间结构,小生境类型多样,生态环境比较脆弱。2011年1月用森林罗盘仪将其划分为20 m × 20 m的方格,进行规则网格法采样,共获得176个样点。样点1 m半径范围内用土钻测定土壤深度3次,取平均值作为土壤厚度;样点3 m半径范围内,采用1 m × 1 m样框测定框内植被所占比例,测定3次,平均值作为植被覆盖度;采样时先去除地表凋落物,在每个样点周围2 m 范围内随机采取5个0—20 cm 表层土样,采集的土样迅速带回实验室,除去土壤中可见的动植物残体,过2 mm 筛,混匀,分为两份。一份置于4 ℃的冷库中,用于土壤MBC、MBN、MBP的测定。测定前将土样预培养7—15 d,消除土壤水分限制对微生物的影响。另一份风干用于土壤理化性质的测定。
1.2.2 样品分析土壤微生物生物量C、N、P 采用氯仿熏蒸提取法测定[13]。MBC采用氯仿熏蒸-K2SO4 提取-碳自动分析法,提取液中C采用总有机碳自动分析仪(TOC2500)测定,MBC=EC/kEC,EC = 熏蒸土壤浸提的有机碳-不熏蒸土壤浸提的有机碳,kEC为转换系数,取值0.45;MBN采用氯仿熏蒸-K2SO4提取-氮自动分析仪法,提取液中N采用流动注射仪(FIAstar 5000)测定,MBN = EN/kEN,EN = 熏蒸土壤浸提的全氮-不熏蒸土壤浸提的全氮,kEN为转换系数,取值0.45;MBP采用氯仿熏蒸-NaHCO3提取-紫外分光光度计法,提取液中P采用紫外分光光度计(UV8500)测定,MNP=EPi/ (kPi·RPi),EPi=熏蒸土壤提取的Pi-不熏蒸土壤提取的Pi,RPi = [(加Pi的土壤提取的Pi-未熏蒸土壤提取的Pi)/25] ×100%,kPi为转换系数,取值0. 4。
1.3 数据处理与统计分析描述性统计分析、典型相关分析在SPSS 13. 0中完成。文中数据采用样本均值加减3倍标准差识别特异值,在此区间外的数据均定为特异值,分别用正常的最大和最小值代替[14],后续计算均采用处理后的原始数据。半方差分析在GS+中完成。Kriging等值线图在ArcGIS10. 0中完成。地统计学有关方法及原理见文献[15] 。
1.3.1 自相关分析空间自相关分析是生态学上常用的空间分析方法,主要用于度量某一变量是否存在空间依赖关系以及变量自身在较近的空间中是否有较强的关联[16]。常用的空间自相关系数有Moran′s I系数和Geary′s C 系数,本文用Moran′s I系数进行空间自相关分析,计算公式为:

式中,xi和xj分别是变量x在相邻配对空间点i和j上取值;wij是相邻权重;n是空间单元总数;I系数取值从-1到1;当I=0时代表空间不相关,取正值时为正相关,取负值为负相关。
1.3.2 半方差函数分析半方差函数是应用最广泛的空间格局描述工具,半方差函数公式为:
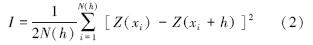
式中,I(h)为半方差函数值;N(h)是间距为向量h的点对总数;Z(xi)是区域化变量Z在xi处的实测值;Z(xi+h)是与xi距离为向量h处样点的值。 一般认为半方差函数只有在最大间隔的1/2内才有意义,在本研究中没有特殊说明半方差函数的有效滞后距都设为其最大采样间隔的1/2。本文对半变异函数的拟合主要采用指数模型(Exponential),其公式为:
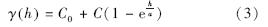
式中,C0为块金值(Nugget);C0+C为基台值(Sill);a是变程(Range);h为滞后距离。一般用R2来衡量模型的好坏。
2 结果与分析 2.1 描述性统计分析通过对偏度、峰度的观察和K-S 法进行非参数检验,在5%的检验水平下,只有MBP服从正态分布。MBC、MBN、MBC/MBN、MBC/MBP经过对数转换后服从正态分布,后续的地统计学分析采用转化后的数据。由表 1可看出,西南峡谷型喀斯特典型坡地的土壤MBC、MBN和MBP比较适中,其均值分别为75.60、25.23和24.06 mg/kg。MBC、MBN、MBP变异系数均远大于75%,呈强度变异,反映了喀斯特生境的高度异质性,内部存在较大变异,但对于其空间结构特征及其相互关系还需用地统计学方法进一步研究。
| 微生物量Microbial biomass | 最小值 Min. /(mg/kg) | 最大值 Max. /(mg/kg) | 均值 Mean /(mg/kg) | 标准差 Standard variance | 变异系数 Variance /% | 偏度 Skew | 峰度 Kurtosis | K-S值 Value of K-S test | 分布类型 Distribution type |
| Cmic:微生物量碳Soil microbial biomass carbon; Cmic#:对数转换后的微生物量碳The Soil microbial biomass carbon after logarithmic; Nmic:微生物量氮Soil microbial biomass nitrogen; Nmic#:对数转换后的微生物量氮The Soil microbial biomass nitrogen after logarithmic; Pmic:微生物量磷Soil microbial biomass phosphorus; Cmic/Nmic:微生物量碳氮比Soil microbial biomass carbon and nitrogen ratio; Cmic/Nmic#:对数转化后的微生物量碳磷比The Soil microbial biomass carbon and nitrogen ratio after logarithmic; Cmic/Pmic:对数转化后的微生物量碳磷比Soil microbial biomass carbon and phosphorus ratio; Cmic/Pmic#:对数转化后的微生物量碳磷比The Soil microbial biomass carbon and phosphorus ratio after logarithmic; #对数转换后的结果; *表示α < 0.05,非正态分布; N表示正态分布 | |||||||||
| Cmic | 13.911 | 161.259 | 75.62 | 33.840 | 114.512 | 0.756 | 0.276 | 0.020 | * |
| Cmic# | 2.633 | 5.08 | 4.22 | 0.483 | 0.233 | -0.604 | 0.693 | 0.169 | N |
| Nmic | 3.201 | 61.011 | 25.23 | 13.519 | 182.756 | 0.940 | 0.286 | 0.007 | * |
| Nmic# | 1.163 | 4.11 | 3.08 | 0.556 | 0.309 | -0.335 | 0.212 | 0.970 | N |
| Pmic | 1.365 | 54.598 | 24.06 | 11.720 | 137.354 | 0.381 | -0.715 | 0.126 | N |
| Cmic/Nmic | 0.61 | 43.45 | 3.87 | 3.776 | 14.193 | 6.525 | 60.861 | 0.000 | * |
| Cmic/Nmic# | -0.493 | 3.77 | 1.14 | 0.618 | 0.382 | 0.415 | 1.388 | 0.892 | N |
| Cmic/Pmic | 0.44 | 34.59 | 4.31 | 3.816 | 14.564 | 3.334 | 19.512 | 0.000 | * |
| Cmic/Pmic# | -0.82 | 3.54 | 1.18 | 0.745 | 0.555 | 0.088 | -0.045 | 0.952 | N |
西南峡谷型喀斯特坡地土壤MBC、MBN、MBP均呈现一定的空间结构分布,三者的差别很大(图 1)。随着滞后距离的增大,MBC呈直线下降,100 m之后呈负相关且继续下降至150 m,在将至坡脚时又有增大的趋势,Moran′s I最小时为-0.0965。MBN随着滞后距的增大呈现下降趋势,在下降至80 m处呈负相关,之后在110 m之后表现为正相关,然后继续下降至180 m,Moran′s I最小为-0.0556。MBP随着滞后距的增大逐渐下降,至60 m时自相关函数逐渐向负方向增长,而到80 m之后又开始增大,在100 m处向负方向增长,之后达到显著的正相关并缓慢增长,在150 m之后又开始急剧下降且在170 m之后达到负相关,Moran′s I最小时为-0.0913;而MBC/MBN的空间结构显著不同于MBC、MBN、MBP的空间结构,随着滞后距的增大开始持续下降并在80 m处下降到最大值,达到最大负相关,之后持续上升,在140处达到显著的正相关并持续上升,Moran′s I最小时为-0.0682;MBC/MBP的空间结构总体趋势是先急剧下降到某一位置再缓慢增大到正相关,然后直线下降,在40 m处急剧下降到负相关,之后60 m处缓慢增长,在120 m处表现为直线下降趋势,Moran′s I最小为-0.0654。

|
| 图 1 土壤微生物量的空间相关图 Fig. 1 Spatial correlograms of soil microbial biomass |
由表 2可看出,西南峡谷型喀斯特坡地MBC、MBN、MBP和MBC/MBN的最佳拟合模型均为指数模型,MBC/MBP的最佳拟合模型为球状模型。除MBP外,RSS均很小,表明拟合程度均很高。土壤微生物量的块金方差C0均很小,接近0,C0/(C0+C)均<25%,说明土壤微生物量均表现为强烈的空间相关,其空间变异受随机因素的影响较小,主要由土壤母质、地形、气候等自然因素(结构性变异)引起。由图 2可看出,MBC/MBN的C0接近于0,表现为纯块金效应。MBC、MBN的C0均小于0. 1,远小于各自的C0+C。MBP的C0较大,可能是试验误差造成的。变程表明属性因子空间自相关范围的大小,它与观测尺度以及在取样尺度上影响土壤微生物的各种生态过程和相互作用有关,在变程之内,变量具有空间自相关性,反之则不存在,即变程提供了研究某种属性相似范围的一种测度[17, 18]。MBN和MBP的变程相近,分别为40.5 m和30.9 m,说明其空间连续性差,空间异质性高。MBC的变程较大,为60.6 m,说明其空间连续性相对较好。
| 微生物量 Microbial biomass | 样本数 Samples | 模型 Theory models | 块金值 C0 | 基台值 C0+C | 块金值/ 基台值 C0/ (C0+C) | 变程 A/m | 拟合优度 R2 | 最小残差值 RSS | 分维数 D |
| Cmic/(mg/kg) | 176 | Exponential | 0.0359 | 0.2418 | 0.149 | 60.6 | 0.839 | 5.465×10-4 | 1.928 |
| Nmic/(mg/kg) | 176 | Exponential | 0.0245 | 0.305 | 0.080 | 40.5 | 0.221 | 4.595×10-3 | 1.939 |
| Pmic/(mg/kg) | 176 | Exponential | 13.4 | 135.7 | 0.099 | 30.9 | 0.510 | 112 | 1.977 |
| Cmic/Nmic | 176 | Exponential | 0.045 | 0.378 | 0.119 | 43.8 | 0.519 | 2.391×10-3 | 1.951 |
| Cmic/Pmic | 176 | Spherical | 0.038 | 0.610 | 0.062 | 33.0 | 0.435 | 3.879×10-3 | 1.971 |

|
| 图 2 土壤微生物量的半方差函数图 Fig. 2 Semivariograms of soil microbial biomass |
利用Kriging方法制作的等值线更深刻、全面和直观地揭示了西南峡谷型喀斯特坡地土壤微生物的空间分布格局(图 3)。与空间自相关和半方差函数分析的结果相似,MBC、MBN、MBP三者的空间分 布格局特征差异很大,表现为极高的空间异质性。

|
| 图 3 土壤微生物量的空间分布 Fig. 3 Spatial distribution of soil microbial biomass |
MBC的高值区集中在坡中上部;MBN表现为上部、坡脚的含量较高,最高值区均分布在坡上部;MBP的格局明显不同,高值区集中在坡脚;MBC/MBN斑块较大,变化缓和;MBC/MBP的空间分布规律不明显,斑块多而破碎,表现为极高的空间异质性。这可能与研究区生境复杂多样、施肥管理及放牧有关。
典型相关分析研究两组变量(两个集团)之间整体的线性相关关系。把5个土壤微生物量(X1—X5)构成第1组变量,把土层厚度、植被覆盖度及土壤理化性状等环境因子(Y1—Y10)构成第2组变量。用典范相关分析来研究土壤微生物与环境因子之间的关系(表 3),并建立典型变量构成(表 4)。前4个特征值的方差累积贡献率达78.00%,基本能反映出大部分变量的信息,由此而建立了土壤微生物量与环境因子之间的4对典型变量构成,由于3、4对典型变量的影响较小,表 4只列出第1、2对典范变量构成进行分析。
| 典型向量 Typical vector | 典型相关系数 Canonical correlation coefficient | 特征值 Eigenvalue | 卡方值 Chi-square value | 自由度 Freedom degree | 显著水平 Significant level | 累积贡献率 Cumulative contribution/% |
| 1 | 0.9966 | 4.3148 | 66.8044 | 50 | 0.0462 | 28.77 |
| 2 | 0.9419 | 3.2135 | 31.8931 | 36 | 0.6643 | 50.19 |
| 3 | 0.8456 | 2.7672 | 16.6151 | 24 | 0.8646 | 68.64 |
| 4 | 0.6937 | 1.4044 | 7.8267 | 14 | 0.8982 | 78.00 |
| M1=-1.3442X1+1.4578X2+0.4926X3+0.1919X4+0.4326X5 |
| X1: 土壤微生物量碳 (MBC); X2: 土壤微生物量氮(MBN); X3: 土壤微生物量磷(MBP); X4: MBC/MBN; X5: MBC/MBP; Y1: 土壤厚度; Y2: 植被覆盖度; Y3: pH; Y4: 有机碳; Y5: 全氮; Y6: 全磷; Y7: 全钾; Y8: 碱解氮; Y9: 速效磷; Y10: 速效钾 |
| M2=0.219X1-0.0482X2+0.0463X3-0.3058X4-1.0539X5 |
| E1=1.0489Y1+0.0065Y2+0.7956Y3-0.1154Y4+0.3483Y5-0.4119Y6+0.3886Y7+0.9401Y8+0.6774Y9+0.0976Y10 |
| E2=-2.0248Y1+0.3859Y2+0.2736Y3-0.3354Y4-0.8925Y5+0.0316Y6-0.2416Y7-1.0547Y8-0.3834Y9+0.009Y10 |
土壤微生物量与环境因子的关系较密切,第1、2对的典型相关系数为0.9419、0.9966,第1对达到了显著水平(<0.05)。土壤微生物量与环境因子的关系中,以MBC、MBN和土层厚度、pH、碱解氮的载荷量较大,说明土层厚度、pH、碱解氮是影响土壤微生物量碳和氮的主要因子。
3 讨论土壤微生物是土壤有机质和养分转化、循环的驱动力,参与有机质的分解、腐殖质的形成等各个生化过程,与土壤肥力密切相关。前期研究已发现喀斯特峰丛洼地景观单元内不同生态系统的植被、土壤养分、水分的空间变异和分布不同,植被、土壤和地形的耦合关系也不同[19, 20, 21],土壤微生物与土壤性质和植被存在着“共演替”效应[10, 11]。本研究中,土壤微生物量在峡谷型喀斯特坡地的植被状况、地形地貌、土壤性质等因子的相互作用下形成了独特的空间格局。
不同土壤类型及生态环境条件下土壤微生物量的变异很大。以往的研究表明,喀斯特峰丛洼地土壤MBC、MBN、MBP的变化范围分别为42—2064、130—216和10—50 mg/kg[22]。李新爱等[11]研究表明,喀斯特区域稻田、林地、旱地土壤Cmic含量主要分布区间分别为550—2550、50—550和50—1050 mg/kg,魏亚伟等人的研究结果表明[23],喀斯特土壤微生物量含量显著高于其它生态系统。本研究中MBC、MBN、MBP的均值分别为75.62、25.23、24.06 mg/kg[24]。MBC与MBP位于该范围内,MBN均值明显低出该范围。峡谷型喀斯特坡地植被、生境的高异质性导致土壤微生物量的变化范围非常广。前人研究中的MBC/MBN均接近于10[25],本研究为4.78,明显低于该值,表明喀斯特生态系统中土壤微生物量氮是植物有效氮的重要储备库。有研究结果表明[26],MBC、MBN、MBP三者之间均呈显著相关,本研究中MBC、MBN呈显著的线性相关关系,与已有的研究结果一致,这可能与研究区域岩石裸露率高导致的采样误差有关。
本研究发现西南喀斯特坡地土壤MBC、MBN、MBP、MBC/MBN和均具有较强的空间自相关性,除MBC/MBP半方差函数的最佳拟合模型为球状模型外,其他指标均为指数模型,RSS均较小,拟合程度较高(半方差函数模型的确定主要依据拟合参数R2和RSS值,这两个参数均是反映模型拟合程度的,但RSS较R2更灵敏,模型拟合RSS值越小,说明拟合程度越高[14];块金值C0较小,C0/(C0+C)均<25%,变程A较短,强烈的空间相关性受随机因素的影响较小,主要由结构性变异引起;由此可见,西南喀斯特坡地土壤微生物生物量在小尺度范围内存在着明显的空间变异和分布,其形成机制与动植物等大型生物一样,是历史进化事件(距离分隔、物理屏障、扩散历史和过去的环境异质性等)和当代环境因子(如植被、气候、地形、土壤和人为干扰等)共同作用的结果[27],且这种影响具有明显的尺度依赖性,即在较大空间尺度的历史及进化过程的主导作用下,当代环境因子在小空间尺度下不断对土壤微生物空间分布格局进行细部改造[28, 29]。
土壤微生物与植物、土壤理化环境之间相互作用、共同演替发展。典范相关分析结果表明,土壤微生物量与环境因子之间存在着较高的相关性,第一、二对典型相关系数在0.9419—0.9966之间,第一对达到了显著水平(<0.05)。西南峡谷型喀斯特坡地土壤微生物量空间分布的影响因子很多,其中土层厚度、pH、碱解氮对土壤微生物量碳(MBC)、氮(MBN)的影响最大,且与MBC呈负相关、与MBN呈正相关。这为指导喀斯特退化生态系统植被的迅速恢复和生态重建、推动微生物生物地理学发展提供了理论基础和实践依据。
| [1] | Doran J W, Zeiss M R. Soil health and sustainability: managing the biotic component of soil quality. Applied Soil Ecology, 2000, 15(1): 3-11. |
| [2] | Zeng F P, Peng W X, Song T Q, Wang K L, Wu H Y, Song X J, Zeng Z X. Changes in vegetation after 22 years′ natural restoration in the karst disturbed area in Northwest Guangxi. Acta Ecologica Sinica, 2000, 27(12): 5110-5119. |
| [3] | Zhou Z X. Scientific Investigation Report of Maolan Karst Forest. Guiyang: Guizhou Science and Technology Press, 1987: 1-23. |
| [4] | Peng Y, Li X Q, Cheng J Z, Xing Y, Yan H. Influence of vegetation types and seasonal variation on soil microbial biomass and microbial respiration in Karst region of Guiyang, Southwest China. Geochimica, 2010, 39(3): 266-273. |
| [5] | Drakare S, Lennon J J, Hillebrand H. The imprint of the geographical, evolutionary and ecological context on species-area relationships. Ecology Letters, 2006, 9(2): 215-227. |
| [6] | Liu S L, Xiao H A, Tong C L, Wu J S. Microbial biomass C, N and P and their responses to application of inorganic and organic fertilizers in subtropical paddy soils. Research of Agricultural Modernization, 2003, 24(4): 278-283. |
| [7] | Feng S Z, Su Y R, Qin X M, Xiao W, Ge Y H, He X Y. Responses of soil microbial properties in soil profile to typical vegetation pattern and slope in karst-cluster depression area. Acta Ecologica Sinica, 2013, 33(10): 3148-3157. |
| [8] | Chen G C, He Z L. The research of microbial under different methods of using on red soil. Chinese Journal of Soil Science, 1998, 29(6): 276-278. |
| [9] | Jin F H, Li S Q, Lu H L, Li S X. Relationships of microbial biomass carbon and nitrogen with particle composition and nitrogen mineralization potential in calcareous soil. Chinese Journal of Applied Ecology, 2007, 18(12): 2739-2746. |
| [10] | He X Y, Wang K L, Chen Z H, Yang G, Chen H S. Responses of soil microbial characters to farm land-use types in peak-cluster depression of Karst region. Chinese Journal of Soil Science, 2008, 39(3): 509-513. |
| [11] | Li X A, Xiao H A, Wu J S, Su Y R, Huang D Y, Huang M, Liu S L, Peng H C. Effects of land use type on soil organic carbon, total nitrogen,and microbial biomass carbon and nitrogen contents in Karst region of South China. Chinese Journal of Applied Ecology, 2006, 17 (10): 1827-1831. |
| [12] | Wu H Y, Zeng F P, Song T Q, Peng W X, Li X H, Ouyang Z W. Spatial variations of soil organic carbon and nitrogen in peak-cluster depression areas of Karst region. Plant Nutrition and Fertilizer Science, 2009, 15(5): 1029-1036. |
| [13] | Wu J S, Lin Q M, Huang Q Y. Soil Microbial Biomass-Methods and Application. Beijing: China Meteorological Press, 2006. |
| [14] | Liu F C, Shi X Z, Yu D S, Pan X Z. Characteristics of spatial variability of total soil nitrogen in the typical area of Taihu Lake basin. Geographical Research, 2004, 23(1): 163-170. |
| [15] | Wang Z Q. Geostatistics and Its Application in Ecology. Beijing: Science press, 1999. |
| [16] | Wang Q, Dai J L, Fu H C, Shen T L, Wu D Q, Wang R Q. The application of spatial analysis methods to microbial ecology. Acta Ecologica Sinica, 2010, 30(2): 439-446. |
| [17] | Cambardella C A, Moorman T B, Parkin T B, Karlen D L, Novak J M, Turco R F, Konopka A E. Field-scale variability of soil properties in Central Iowa soils. Soil Science Society of America Journal, 1994, 58(5): 1501-1511. |
| [18] | Trangmar B B, Yost R S, Uehara G. Application of geostatistics to spatial studies of soil properties. Advances in Agronomy, 1986, 38: 45-94. |
| [19] | Song T Q, Peng W X, Zeng F P, Ouyang Z W, Wu H Y. Spatial heterogeneity of surface soil moisture content in dry season in Mulun National Natural Reserve in Karst area. Chinese Journal of Applied Ecology, 2009, 20(1): 98-104. |
| [20] | Peng W X, Wang K L, Song T Q, Zeng F P, Wang J R. Controlling and restoration models of complex degradation of vulnerable Karst ecosystem. Acta Ecologica Sinica, 2008, 28(2): 811-820. |
| [21] | Du H, Song T Q, Peng W X, Wang K L, Liu L, Lu S Y, Zeng F P. Spatial heterogeneity of mineral compositions in surface soil in Mulun National Nature reserve Karst areas. Transactions of the Chinese Society of Agricultural Engineering, 2011, 27(6): 79-84. |
| [22] | Zhang L Q, Peng W X, Song T Q, Zou D S, Zeng F P, Song M, Yu Z, Liu Y. Spatial heterogeneity of soil microbial biomass carbon, nitrogen, and phosphorus in sloping farmland in a karst region on the Yunnan-Guizhou Plateau. Acta Ecologica Sinica, 2012, 32(7): 2056-2065. |
| [23] | Wei Y W, Su Y R, Chen X B, He X Y. Effects of human disturbance on profile distribution of soil organic C,total N, total P and microbial biomass in Karst region of Northwest Guangxi. Journal of Soil and Water Conservation, 2010, 24(3): 164-169. |
| [24] | Liu S, Wang C K. Spatial-temporal patterns of soil microbial biomass carbon and nitrogen in five temperate forest ecosystems. Acta Ecologica Sinica, 2010, 30(12): 3135-3143. |
| [25] | Wu J G, Ai L. Soil microbial activity and biomass C and N content in three typical ecosystems in Qilian Mountains, China. Journal of Plant Ecology, 2008, 32(2): 465-476. |
| [26] | Xue S, Liu G B, Dai Q H, Li X L, Wu R J. Dynamic changes of soil microbial biomass in the restoration process of shrub plantations in loess hilly area. Chinese Journal of Applied Ecology, 2008, 19(3): 517-523. |
| [27] | Jin Z Z, Lei J Q, Xu X W, Li S Y, Fan J L, Zhao S F, Zhou H W, Gu F. Relationships of soil microbial biomass with soil environmental factors in Tarim Desert highway shelter-forest. Chinese Journal of Applied Ecology, 2009, 20(1): 51-57. |
| [28] | Zhang W, Chen H S, Wang K L, Hou Y, Zhang J G. Spatial variability of soil organic carbon and available phosphorus in a typical Karst depression, northwest of Guangxi. Acta Ecologica Sinica, 2007, 27(12): 5168-5175. |
| [29] | Cheng Y, An S S, Li G H, Li D H. Soil nutrient and microbial biomass in the Loess hilly area of Ningxia under different plant rehabilitation patterns. Chinese Journal of Eco-Agriculture, 2010, 18(2): 261-266. |
| [2] | 曾馥平, 彭晚霞, 宋同清, 王克林, 吴海勇, 宋希娟, 曾昭霞. 桂西北喀斯特人为干扰区植被自然恢复22年后群落特征. 生态学报, 2007, 27(12): 5110-5119. |
| [4] | 彭艳, 李心清, 程建中, 邢英, 闫慧. 贵阳喀斯特地区植被类型与季节变化对土壤微生物生物量和微生物呼吸的影响. 地球化学, 2010, 39(3): 266-273. |
| [6] | 刘守龙, 肖和艾, 童成立, 吴金水. 亚热带稻田土壤微生物生物量碳、氮、磷状况及其对施肥的反应特点. 农业现代化研究, 2003, 24(4): 278-283. |
| [7] | 冯书珍, 苏以荣, 秦新民, 肖伟, 葛云辉, 何寻阳. 喀斯特峰丛洼地土壤剖面微生物特性对植被和坡位的响应. 生态学报, 2013, 33(10): 3148-3157. |
| [8] | 陈国潮, 何振立. 红壤不同利用方式下的微生物量研究. 土壤通报, 1998, 29(6): 276-278. |
| [9] | 金发会, 李世清, 卢红玲, 李生秀. 石灰性土壤微生物量碳、氮与土壤颗粒组成和氮矿化势的关系. 应用生态学报, 2007, 18(12): 2739-2746. |
| [10] | 何寻阳, 王克林, 于一尊, 张伟, 陈志辉. 岩溶区植被和季节对土壤微生物遗传多样性的影响. 生态学报, 2009, 29(4): 1763-1769. |
| [11] | 李新爱, 肖和艾, 吴金水, 苏以荣, 黄道友, 黄敏, 刘守龙, 彭洪翠. 喀斯特地区不同土地利用方式对土壤有机碳、全氮以及微生物生物量碳和氮的影响. 应用生态学报, 2006, 17(10): 1827-1831. |
| [12] | 吴海勇, 曾馥平, 宋同清, 彭晚霞, 黎星辉, 欧阳资文. 喀斯特峰丛洼地土壤有机碳和氮素空间变异特征.植物营养与肥料学报, 2009, 15(5): 1029-1036. |
| [13] | 吴金水, 林启美, 黄巧云. 土壤微生物生物量测定方法及其应用. 北京: 气象出版社, 2006. |
| [14] | 刘付程, 史学正, 于东升, 潘贤章. 太湖流域典型地区土壤全氮的空间变异特征. 地理研究, 2004, 23(1): 163-170. |
| [15] | 王政权. 地统计学及在生态学中的应用. 北京: 科学出版社, 1999. |
| [16] | 王强, 戴九兰, 付合才, 申天琳, 吴大千, 王仁卿. 空间分析方法在微生物生态学研究中的应用. 生态学报, 2010, 30(2): 439-446. |
| [19] | 宋同清, 彭晚霞, 曾馥平, 欧阳资文, 吴海勇. 喀斯特木论自然保护区旱季土壤水分的空间异质性. 应用生态学报, 2009, 20(1): 98-104. |
| [20] | 彭晚霞, 宋同清, 曾馥平, 王克林, 傅伟, 刘璐, 杜虎, 鹿士杨, 殷庆仓. 喀斯特常绿落叶阔叶混交林植物与土壤地形因子的耦合关系. 生态学报, 2010, 30(13): 3472-3481. |
| [21] | 杜虎, 宋同清, 彭晚霞, 王克林, 刘璐, 鹿士杨, 曾馥平. 木论喀斯特自然保护区表层土壤矿物质的空间异质性. 农业工程学报, 2011, 27(6): 79-84. |
| [22] | 张利青, 彭晚霞, 宋同清, 邹冬生, 曾馥平, 宋敏, 俞孜, 刘艳.云贵高原喀斯特坡耕地土壤微生物量 C、N、P空间分布.生态学报, 2012, 32(7): 2056-2065. |
| [23] | 魏亚伟, 苏以荣, 陈香碧, 何寻阳. 人为干扰对桂西北喀斯特生态系统土壤有机碳、氮、磷和微生物量剖面分布的影响. 水土保持学报, 2010, 24(3): 164-169. |
| [24] | 刘爽, 王传宽. 五种温带森林土壤微生物生物量碳氮的时空格局. 生态学报, 2010, 30(12): 3135-3143. |
| [25] | 吴建国, 艾丽. 祁连山3种典型生态系统土壤微生物活性和生物量碳氮含量. 植物生态学报, 2008, 32(2): 465-476. |
| [26] | 薛箑, 刘国彬, 戴全厚, 李小利, 吴瑞俊. 黄土丘陵区人工灌木林恢复过程中的土壤微生物生物量演变. 应用生态学报, 2008, 19(3): 517-523. |
| [27] | 靳正忠, 雷加强, 徐新文, 李生宇, 范敬龙, 赵思峰, 周宏伟, 谷峰. 塔里木沙漠公路防护林土壤微生物生物量与土壤环境因子的关系. 应用生态学报, 2009, 20(1): 51-57. |
| [28] | 张伟, 陈洪松, 王克林, 侯娅, 张继光. 桂西北喀斯特洼地土壤有机碳和速效磷的空间变异. 生态学报, 2007, 27(12): 5168-5175. |
| [29] | 成毅, 安韶山, 李国辉, 李第红. 宁夏黄土丘陵区植被恢复对土壤养分和微生物生物量的影响. 中国生态农业学报, 2010, 18(2): 261-266. |