 2014, Vol. 34
2014, Vol. 34文章信息
- 易好, 邓湘雯, 项文化, 方晰, 赵丽娟, 李家湘, 崔飞
- YI Hao, DENG Xiangwen, XIANG Wenhua, FANG Xi, ZHAO Lijuan, LI Jiaxiang, CUI Fei
- 湘中丘陵区南酸枣阔叶林群落特征及群落更新
- The characteristics and regeneration of the Choerospondias axillaries broad-leaved community in the hilly region of central Hunan Province, China
- 生态学报, 2014, 34(12): 3463-3471
- Acta Ecologica Sinica, 2014, 34(12): 3463-3471
- http://dx.doi.org/10.5846/stxb201309302403
-
文章历史
- 收稿日期:2013-09-30
- 修订日期:2014-04-04






2. 中南林业科技大学林学院, 长沙 410004
2. Faculty of Forestry, Central South University of Forestry and Technology, Changsha 410004, China
落叶阔叶林是分布于我国暖温带地区的主要地带性植被[1]。我国学者对温带地区的落叶阔叶林的区系组成、群落结构和演替过程进行了很多研究,例如:有学者发现我国暖温带落叶阔叶林多以栎林为主[2]。后来有人采用样地调查的方法对辽东栎(Quercus liaotungensis)的种群结构和动态进行了研究[3],并提出了利用种间联结性来保护植物群落稳定性的方法[4],这些研究成果有效的应用到了保护我国的落叶阔叶林资源的工作中。在我国亚热带地区也分布有不少的落叶阔叶林[5],且这些地区分布的落叶阔叶林的生物多样性要高于暖温带[6],然而有学者发现我国亚热带地区的落叶阔叶林正在衰退[7],亚热带阔叶林的研究工作逐渐引起重视[8, 9],目前对如何保护和利用亚热带地区的落叶林的研究还不多[10]。
南酸枣阔叶林是分布在我国长江以南的亚热带地区常见的次生林群落类型之一[11]。南酸枣是漆树科、漆树属的植物,为中性偏阳、生长快、适应性强的落叶乔木[12],也是我国南方的常见的阔叶林群落类型,在南方森林群落中占有比较重要的地位[13],国内对南酸枣的研究主要集中在其经济价值和药用价值上[14],针对其生态结构与功能的研究很少。
湘中丘陵区是隶属于亚热带地区的重要组成之一,2006年,魏勇等人在长沙县大山冲森林公园通过线路调查的方式进行植被调查,发现南酸枣的幼苗数量明显少于其他树种,并指出南酸枣阔叶林群落处于衰退期[15],但是其研究并没有用具体的数据说明群落的结构及当前的生长状态。本研究通过对长沙县大山冲森林公园内的南酸枣阔叶林植物群落的结构特征进行调查,研究湘中丘陵区南酸枣阔叶林群落结构的组成及更新状况,探讨南酸枣优势种群的分布格局,为南酸枣阔叶林的保护、生产经营和生态功能的研究提供理论依据,为认识南酸枣群落的发生与演替提供数据支撑,为湘中丘陵区生态系统的恢复与重建提供指导。
1 研究区概况研究地位于湖南省长沙县大山冲林场内,地理坐标为28°23′58″—28°24′58″N,113°17′46″—113°19′08″E,地处幕阜山余脉的西缘,海拔55—217.4m,属湘中丘陵地貌。气候为中亚热带东南季风湿润气候区,年平均气温16.6—17.6℃,极端最高温40℃,最低温-11℃,年平均日照1300—1800h,无霜期345d,年平均降水量1412—1559mm[16]。土壤为板页岩发育而成的红壤,地带性植被为亚热带常绿阔叶林。由于过去人为干扰,其原生植被大都受到不同程度的破坏[17],现存的植被类型主要有杉木人工林、马尾松次生林、南酸枣林、石栎-青冈林等,其中南酸枣阔叶林为林场内保存较为完好的阔叶林之一。
2 研究方法 2.1 样地设置与调查2013年4月在长沙县大山冲森林公园内设置1hm2的南酸枣阔叶林固定样地,并在样地内设置10m×10m样方100个,每个样方的边界用绳子分隔,呈网格状。
对100个样方中胸径大于1cm的所有植物进行详细调查,记录其种名、胸径、树高、第1活枝高、冠幅、生活力等,并进行每木定位(确定x,y坐标)。在大样地中根据不同的坡位和坡向共选取了16个2m×2m小样方调查胸径小于1cm的幼苗,记录其种名、数量。
2.2 数据统计与分析本研究主要采用重要值作为种在群落中的重要程度参数和指标,其计算公式如下[18]:

式中,Itj为乔木重要值。

式中,Ihsj为灌木草本重要值。
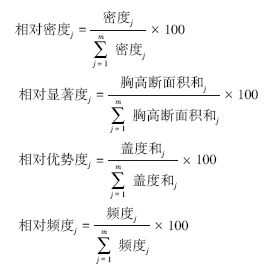
数据采用EXCEL 2007软件进行汇总分析;
乔木层成层结构的划分:将乔木层按树高划分为5个林层,第1层的树高范围为H>25m,第2层的树高范围为15m<H≤25m,第3层的树高范围为10m<H≤15m,第4层的树高范围为5m<H≤10m,第5层树高范围H≤5m[19]。
3 结果与分析 3.1 植物种类组成根据野外调查记录的100个样方的统计,南酸枣阔叶林中胸径大于1cm的树种有59个种,隶属于42属25科(表 1)。其中乔木41种,灌木18种。常绿树种25个,落叶树种34个。主要的科属有壳斗科4属7种,樟科4属6种,山茶科4属5 种,山矾科1属5种,蔷薇科4属5种。
| 科 Family | 属 Genus | 种 Species |
| 壳斗科 Fagaceae | 柯属 Lithocarpus | 米槠Castanopsis carlesii |
| 锥属 Castanopsis | 甜储Castanopsis eyrei,锥栗Castanea henryi | |
| 栎属 Quercus | 石栎Lithocarpus glaber,白栎Quercus fabri,槲栎Quercus aliena, 青冈Cyclobalanopsis glauca | |
| 樟科 Lauraceae | 檫木属 Sassafras | 檫木Sassafras tzumu |
| 樟属 Cinnamomum | 樟树Cinnamomum camphora | |
| 山胡椒属 Lindera | 山胡椒Lindera angustifolia,山橿Lindera reflexa, | |
| 木姜子属 Litsea | 毛豹皮樟Litsea coreana,山鸡椒Litsea cubeba | |
| 山茶科 Theaceae | 红淡比属 Cleyera | 红淡比Cleyera japonica |
| 木荷属 Schima | 木荷Schima superba | |
| 山茶属 Camellia | 油茶Camellia oleifera | |
| 柃木属 Eurya | 格药柃Eurya muricata,细枝柃Eurya loquaiana | |
| 山矾科 Symplocaceae | 山矾属 Symplocos | 白檀Symplocos paniculata,老鼠矢Symplocos stellaris,南岭山矾Symplocos confusa,山矾Symplocos caudata |
| 四川山矾Symplocos setchuensis | ||
| 蔷薇科 Rosaceae | 花楸属 Sorbus | 石灰花楸Sorbus folgneri |
| 悬钩子属 Rubus | 山莓Rubus corchorifolius | |
| 樱属 Cerasus | 樱桃Prunus pseudocerasus | |
| 石楠属 Photinia | 椤木石楠Photinia davidsoniae,中华石楠Photinia beauverdiana | |
| 大戟科 Euphorbiaceae | 野桐属 Mallotus | 野桐Mallotus apelta |
| 油桐属 Vernicia | 千年桐Vernicia montana,油桐Vernicia fordii | |
| 冬青科 Aquifoliaceae | 冬青属Ilex | 冬青Ilex purpurea,榕叶冬青Ilex ficoidea,台湾冬青Ilex formosana |
| 杜鹃花科 Ericaceae | 越橘属 Vaccinium | 乌饭Vaccinium bracteatum |
| 杜鹃属 Rhododendron | 满山红Rhododendron mariesii,映山红Rhododendron simsii | |
| 忍冬科 Caprifoliaceae | 荚蒾属 Viburnum | 荚蒾Viburnum dilatatum,南方荚蒾Viburnum fordiae |
| 金缕梅科 Hamamelidaceae | 枫香树属 Liquidambar | 枫香Liquidambar formosana |
| 檵木属 Loropetalum | 檵木Loropetalum chinensis | |
| 柿科 Ebenaceae | 柿属 Diospyros | 延平柿 Diospyros tsangii,野柿 Diospyros kaki |
| 榆科 Ulmaceae | 榉属 Zelkova | 榉树 Zelkova serrata |
| 朴属 Celtis | 朴树 Celtis sinensis | |
| 马鞭草科 Verbenaceae | 大青属 Clerodendrum | 大青 Clerodendrum cyrtophyllum |
| 紫珠属 Callicarpa | 紫珠 Callicarpa bodinieri | |
| 松科 Pinaceae | 松属 Pinus | 马尾松 Pinus massoniana |
| 桃金娘科 Myrtaceae | 蒲桃属 Syzygium | 赤楠 Syzygium buxifolium |
| 省沽油科 Staphyleaceae | 野鸦椿属 Euscaphis | 野鸭椿 Euscaphis japonica |
| 棕榈科 Palmae | 棕榈属 Trachycarpus | 棕榈 Trachycarpus fortunei |
| 桑科 Moraceae | 榕属 Ficus | 异叶榕 Ficus heteromorpha |
| 醉鱼草科 Buddlejaceae | 醉鱼草属 Buddleja | 醉鱼草 Buddleja lindleyana |
| 桤叶树科 Clethraceae | 桤叶树属 Clethra | 恺叶树 Clethra faberi |
| 漆树科 Anacardiaceae | 南酸枣属 Choerospondias | 南酸枣 Choerospondias axillaris |
| 茜草科 Rubiaceae | 栀子属 Gardenia | 栀子 Gardenia jasminoides |
| 杜英科 Elaeocarpaceae | 杜英属 Elaeocarpus | 日本杜英 Elaeocarpus japonicus |
| 八角枫科 Alangiaceae | 八角枫属 Alangium | 毛八角枫 Alangium kurzii |
| 楝科 Meliaceae | 楝属 Melia | 苦楝 Melia azedarach |
南酸枣群落中个乔木树种的重要值计算结果表明(表 2):南酸枣的重要值最高,为23.57,远远高于四川山矾(10.46),在群落中处于优势地位,为群落建群种。四川山矾和格药柃分别排重要值的第2位( 10.46)和第3位(7.98)。
|
种名
Species | 频度/%
Frequency | 株数
Number of plant | 相对密度
Relative density | 相对频度
Relative frequency | 相对显著度
Relative prominence | 重要值
Importance value |
| *表中列出重要值前20位的乔木树种 | ||||||
| 南酸枣Choerospondias axillaris | 80 | 215 | 6.92 | 8.26 | 55.52 | 23.57 |
| 四川山矾Symplocos setchuensis | 70 | 613 | 19.72 | 7.22 | 4.44 | 10.46 |
| 格药柃Eurya muricata | 73 | 453 | 14.57 | 7.53 | 1.85 | 7.98 |
| 千年桐Vernicia montana | 62 | 137 | 4.41 | 6.40 | 5.37 | 5.39 |
| 毛豹皮樟Litsea coreana | 53 | 136 | 4.37 | 5.47 | 2.66 | 4.17 |
| 红淡比Cleyera japonica | 44 | 172 | 5.53 | 4.54 | 1.02 | 3.70 |
| 油桐Vernicia fordii | 45 | 79 | 2.54 | 4.64 | 3.65 | 3.61 |
| 油茶Camellia oleifera | 31 | 177 | 5.69 | 3.20 | 1.18 | 3.36 |
| 榕叶冬青Ilex ficoidea | 42 | 127 | 4.08 | 4.33 | 0.94 | 3.12 |
| 中华石楠Photinia beauverdiana | 45 | 99 | 3.18 | 4.64 | 0.94 | 2.92 |
| 老鼠矢Symplocos stellaris | 40 | 118 | 3.80 | 4.13 | 0.47 | 2.80 |
| 樟树Cinnamomum camphora | 14 | 39 | 1.25 | 1.44 | 5.55 | 2.75 |
| 山矾Symplocos caudata | 42 | 85 | 2.73 | 4.33 | 0.53 | 2.53 |
| 白栎Quercus fabri | 23 | 29 | 0.93 | 2.37 | 3.81 | 2.37 |
| 山鸡椒Litsea cubeba | 35 | 88 | 2.83 | 3.61 | 0.33 | 2.26 |
| 冬青Ilex purpurea | 31 | 79 | 2.54 | 3.20 | 1.02 | 2.25 |
| 青冈Cyclobalanopsis glauca | 27 | 70 | 2.25 | 2.79 | 1.50 | 2.18 |
| 椤木石楠Photinia davidsoniae | 33 | 80 | 2.57 | 3.41 | 0.55 | 2.18 |
| 山胡椒Greyblue Spicebush | 16 | 59 | 1.90 | 1.65 | 0.30 | 1.28 |
| 野柿Diospyros kaki | 15 | 30 | 0.96 | 1.55 | 1.09 | 1.20 |
| 其他 Others | 6.37 | 13.31 | 6.86 | 8.85 |
林下植被有灌木植物23种,草本植物9种,蕨类植物5种。灌木层和草本层中檵木的重要值最大,为54.59(表 3),与其他灌木相比较,它重要值的各个指标都处于绝对优势。满山红在灌木层中的重要值为21.85,在灌木层中的重要值排第二位,其物种数量在群落中的比例很大。
|
种名
Species | 频度/%
Frequency | 株数
Number of plant | 相对密度
Relative density | 相对频度
Relative frequency | 相对优势度
Relative dominance | 重要值
Importance value |
| 檵木Loropetalum chinensis | 95 | 1330 | 60.54 | 32.87 | 70.35 | 54.59 |
| 满山红Rhododendron mariesii | 58 | 568 | 25.85 | 20.07 | 19.63 | 21.85 |
| 乌饭Vaccinium bracteatum | 30 | 56 | 2.55 | 10.38 | 2.02 | 4.98 |
| 栀子Gardenia jasminoides | 29 | 51 | 2.32 | 10.03 | 1.53 | 4.63 |
| 映山红Rhododendron simsii | 18 | 93 | 4.23 | 6.23 | 3.12 | 4.53 |
| 大青Cleredendrumcwtophy | 20 | 39 | 1.78 | 6.92 | 1.41 | 3.37 |
| 异叶榕Ficus heteromorpha | 10 | 13 | 0.59 | 3.46 | 0.34 | 1.47 |
| 赤楠Syzygium buxifolium | 8 | 10 | 0.46 | 2.77 | 0.37 | 1.20 |
| 紫珠Callicarpa bodinieri | 5 | 10 | 0.46 | 1.73 | 0.35 | 0.84 |
| 野桐Mallotus apelta | 4 | 10 | 0.46 | 1.38 | 0.22 | 0.68 |
| 醉鱼草Buddleja lindleyana | 2 | 3 | 0.14 | 0.69 | 0.08 | 0.30 |
| 细枝柃Eurya loquaiana | 2 | 3 | 0.14 | 0.69 | 0.08 | 0.30 |
| 山橿Lindera reflexa | 2 | 2 | 0.09 | 0.69 | 0.08 | 0.29 |
| 荚蒾Viburnum dilatatum | 2 | 2 | 0.09 | 0.69 | 0.05 | 0.28 |
| 恺叶树Clethra faberi | 1 | 3 | 0.14 | 0.35 | 0.25 | 0.24 |
| 白檀Symplocos paniculata | 1 | 2 | 0.09 | 0.35 | 0.08 | 0.17 |
| 南方荚蒾Viburnum fordiae | 1 | 1 | 0.05 | 0.35 | 0.03 | 0.14 |
| 山莓Rubus corchorifolius | 1 | 1 | 0.05 | 0.35 | 0.03 | 0.14 |
径级分布不仅反映了群落的结构特征,也有助于了解群落的动态趋势。从群落的乔木胸径分布的统计结果来看(图 1),径级结构呈倒J型分布,小径级林木(胸径小于6cm)占50%,中径级林木(胸径6—20cm)占33.85%,较大径级林木(胸径大于20cm)占16.15%。统计结果说明群落结构的主要组成部分还是由胸径小于12cm的小乔木种群组成。
为了解乔木层胸径结构呈倒J型分布的具体原因,分别对乔木层重要值最大的5个树种进行径级结构的分析(图 2)。

|
| 图 1 乔木树种的径级分布 Fig.1 Distribution of diameter classes for the canopy tree species |

|
| 图 2 主要乔木树种径级结构分布 Fig.2 Diameter class distribution of several dominant tree species |
作为大型落叶乔木,南酸枣种群数量明显少于格药柃、四川山矾等小乔木(图 2)。南酸枣的径级结构呈正态分布的特征,径级最大的超过54cm,大部分为大径木,属于成熟或近熟的林木,群落中的大径木几乎都由南酸枣组成,其在群落中的优势地位很明显。其径级小于12cm的个体数23株,占种群总数的12.9%,在种群中的比例很小、数量不足。
四川山矾[20]是群落的常见种(图 2),其胸径结构呈倒J型分布,胸径最大为14cm。四川山矾胸径小于4cm的个体幼树占种群的比例为41.1%,四川山矾的数量随胸径结构的增大呈递减趋势,就种群数量而言,它在当前群落发展阶段具有的竞争力最大。
格药柃和四川山矾同为小乔木,都是湘中丘陵地区群落的常见种,其数量少于四川山矾,两个树种在胸径结构上的分布很相似。千年桐也是该群落中常见的落叶阔叶乔木,其胸径大于12cm的个体在各个径级均保持一定的数量(图 2),胸径小于12cm的个体数量有明显逐级增加的趋势,且这一趋势在8cm以下的幼树中表现尤为突出。这种现象的原因是千年桐具有很好的抗寒性[21],因此其幼树在2008年的冰雪灾害中保存良好,主要表现是个体数量的增加。毛豹皮樟喜欢生长于温暖、湿润的亚热带环境,抗逆性强、适应性广[22],是该区域内常见的一种樟科常绿乔木,其种群数量与千年桐相近,胸径的分布趋势表明其在群落未来的发展中将有很强的竞争力。
将乔木层按树高划分为5个林层(表 4),第1层中只有南酸枣、檫木、枫香等3个落叶树种,其中南酸枣10株,占83.3%。第2层的主要组成树种是南酸枣,占69.8%,远大于千年桐(5.0%)、毛豹皮樟(1.4%)、油桐(7.2%)、樟树(3.6%)、白栎(2.9%)、青冈(0.7%)和野柿(0.7%)等树种。第3层,南酸枣植株数量所占百分比为43.5%,千年桐和樟树植株占的百分比分别为14.8%和9.3%。第4层中四川山矾占18.1%,常绿树种在第4层开始占主体,并且以常绿小乔木为主。第5层的优势树种为四川山矾(22.6%),其次是油茶(10.7%)和格药柃(10.4%)。
在前3层中,南酸枣重要值最大且占有极大的比例(83.3%、69.8%、43.5%),充分的体现了大乔木的优势,在较高的林层中,南酸枣数量占绝大多数。在第2、3层中南酸枣的比例相对第1层而言逐渐减少,与图 2中南酸枣中所呈现正态分布的径级结构相照应。在第4层中,南酸枣数量和比例相对第2层和第3层而言急剧减少,毛豹皮樟、四川山矾和千年桐开始大量出现,并且成为这一林层的主要树种。第5层的树种数量最多,乔木幼树、小乔木和灌木等大量物种都是处于这一林层,种间竞争强度大,四川山矾、格药柃、油茶、檵木等在物种数量方面有很大优势。
|
种名
Species | 第1层 First layer
(H>25m) | 第2层 Second layer
(H:16—25m) | 第3层 Third layer
(H:11—15m) | 第4层 Fourth layer
(H:6—10m) | 第5层 Fifth layer
(H:2—5m) | ||||||
| 株数
Numbers | 比例/%
Proportion |
株数
Numbers | 比例/%
Proportion |
株数
Numbers | 比例/%
Proportion |
株数
Numbers | 比例/%
Proportion |
株数
Numbers | 比例/%
Proportion | ||
| *表中列取重要值前20位乔木树种 | |||||||||||
| 南酸枣Choerospondias axillaris | 10 | 83.3 | 97 | 69.8 | 47 | 43.5 | 26 | 6.7 | 6 | 1.8 | |
| 四川山矾Symplocos setchuensis | 70 | 18.1 | 74 | 22.6 | |||||||
| 格药柃Eurya muricata | 1 | 0.9 | 11 | 2.8 | 34 | 10.4 | |||||
| 千年桐Vernicia montana | 7 | 5.0 | 16 | 14.8 | 32 | 8.3 | 2 | 0.6 | |||
| 毛豹皮樟Litsea coreana | 2 | 1.4 | 5 | 4.6 | 37 | 9.6 | 20 | 6.1 | |||
| 红淡比Cleyera japonica | 8 | 2.1 | 25 | 7.6 | |||||||
| 油桐Vernicia fordii | 10 | 7.2 | 6 | 5.6 | 21 | 5.4 | 7 | 2.1 | |||
| 油茶Camellia oleifera | 17 | 4.4 | 35 | 10.7 | |||||||
| 榕叶冬青Ilex ficoidea | 1 | 0.9 | 11 | 2.8 | 24 | 7.3 | |||||
| 中华石楠Photinia beauverdiana | 1 | 0.9 | 20 | 5.2 | 1 | 0.3 | |||||
| 老鼠矢Symplocos stellaris | 5 | 1.3 | 8 | 2.4 | |||||||
| 樟树Cinnamomum camphora | 5 | 3.6 | 10 | 9.3 | 15 | 3.9 | 3 | 0.9 | |||
| 山矾Symplocos caudata | 8 | 2.1 | 13 | 4.0 | |||||||
| 白栎Quercus fabri | 4 | 2.9 | 8 | 7.4 | 10 | 2.6 | 4 | 1.2 | |||
| 山鸡椒Litsea cubeba | 9 | 2.3 | 1 | 0.3 | |||||||
| 冬青Ilex purpurea | 1 | 0.9 | 12 | 3.1 | 12 | 3.7 | |||||
| 青冈Cyclobalanopsis glauca | 1 | 0.7 | 1 | 0.9 | 12 | 3.1 | 12 | 3.7 | |||
| 椤木石楠Photinia davidsoniae | 1 | 0.9 | 8 | 2.1 | 8 | 2.4 | |||||
| 山胡椒Greyblue Spicebush | 8 | 2.1 | 2 | 0.6 | |||||||
| 野柿Diospyros kaki | 1 | 0.7 | 2 | 1.9 | 6 | 1.6 | 4 | 1.2 | |||
| 其它树种 Other species | 2 | 16.7 | 12 | 8.6 | 8 | 7.4 | 40 | 10.4 | 32 | 9.8 | |
| 合计Total | 12 | 100 | 139 | 100 | 108 | 100 | 386 | 100 | 327 | 100 | |
群落的更新是群落演替和发展的重要基础[23],通过对南酸枣阔叶林乔木物种幼苗(1.0cm≤DBH<1.5cm,树高H<1.3m)数量的统计,在1hm2固定样地中,乔木更新的幼苗总数为522株,其中乔木树种南酸枣只有幼苗6株(图 3),仅占幼苗总数的1.1%,南酸枣的幼苗数量严重不足,这将影响南酸枣在未来的竞争,使其难以保持在群落中的优势。四川山矾的幼苗数最多,有135株,占25.9%,其次为格药柃,有76株,占14.6%,这两种小型乔木的幼苗数量之和占群落中幼苗总数的40.5%,极大的幼苗数量让其在未来的竞争中处于优势地位,也表明在下一个演替阶段中它们将占有很重要的地位。千年桐和毛豹皮樟这两种乔木的幼苗数量较少,分别为16株和

|
| 图 3 主要乔木幼树的更新情况 Fig.3 Quantitative characteristics of regeneration of the dominant tree species saplings |
1 5株,结合两个物种在图 2中的胸径结构来看,相对于南酸枣优势度的逐渐降低,两树种的优势度呈现上升的趋势,这种情况对于群落的稳定发展有积极的影响。
就群落当前情况而言,南酸枣作为群落的优势种,幼苗的增补率很小,退化的现象明显,已经表现出了隐退种的特征,优势地位在逐步的下降。
4 讨论与建议 4.1 讨论群落乔木树种中南酸枣重要值最大,主要原因是具有较大的相对显著度,这一特征在径级结构和垂直结构中表现十分明显,在群落中,南酸枣具有极大的优势度,是群落的建群种。乔木组成除优势种南酸枣外还有千年桐和毛豹皮樟等大乔木,这两个树种在群落中保留有一定的数量,但显著度不如南酸枣明显。小乔木以四川山矾和格药柃等常绿阔叶树种为主,两种小乔木主要优势是数量大,朱晓文利用TWINSPAN将湘中丘陵区的岳麓山风景名胜区的6个植被类型划分为18植物群系,其中有11个植物群系都将四川山矾和格药柃确定为植物群系的优势种或伴生种[24],本文的研究结果符合其对湘中丘陵区植物群系特征研究的结果。灌木层的物种以金缕梅科的檵木为主导地位,其次是杜鹃花科的满山红,这两个物种的优势表现在数量和种群分布上。
通过对群落中各物种胸径结构、成层结构以及幼苗更新情况的分析,南酸枣的胸径结构表现出隐退种的特征,幼树和幼苗的数量无论是相对于种群数量或者相对其他竞争种的数量来说都严重不足,这将影响南酸枣在接下来的演替过程中优势种的地位,本研究通过数据分析进一步证明了魏勇所指出的湘中丘陵区南酸枣群落的衰退现象。四川山矾、格药柃、千年桐、毛豹皮樟等几个树种的胸径分布规律和良好的更新情况表明湘中丘陵区的南酸枣阔叶林群落将会朝向常绿阔叶林方向进行演替。根据张净等人的研究,林窗可能是造成南酸枣更新不良的原因,林窗形成后,直接改变生境内的光照、水分和温度等因子,导致土壤理化性质、营养元素分解和土壤微生物活性等发生改变,影响群落内生物学过程的变化[25],并影响了南酸枣幼苗的自然更新发生改变,就南酸枣目前的更新情况而言,这种改变是不利的。2008年的冰雪灾害造成大量的南酸枣植株断干和断梢,这是南酸枣群落内形成林窗的主要原因。
研究结果明显表明南酸枣在群落中呈退化趋势,常绿阔叶树种的优势地位上升,这与曾思齐[11],金则新[7]等人的研究结果一致。特别是在南酸枣群落经过冰雪灾害的破坏以后,南酸枣种群大径级植株遭到破坏,更新情况不良,其退化趋势更加明显。
4.2 建议封山育林的办法能有效避免人类活动对当地生物多样性的直接影响,但是不能阻止自然状态下群落内部物种间的优胜劣汰。鉴于南酸枣群落的现状,保护南酸枣种群在群落的优势地位,也为保存南酸枣群落这一散落于亚热带地区的生态景观,建议相关部门采取措施以保护南酸枣种群在群落中的优势地位,根据本研究的南酸枣径级结构(图 2),可以在群落中补植2—12cm的南酸枣植株,以改善南酸枣种群的径级结构的分布格局。特别是在群落中由于南酸枣断梢断干所形成的林窗区域,补植10—12cm的较大植株,增加郁闭度,以改变林下的温度和光照条件。其次,通过实验从土壤、水分等方面分析南酸枣更新不良的具体原因,通过改变土壤理化性质的方式促进南酸枣的更新,以加强湘中丘陵区南酸枣阔叶林生态系统的恢复与重建工作。
| [1] | Chinese Vegetation Editorial Committee. Chinese Vegetation. Beijing: Science Press, 1980. |
| [2] | Chen L Z, Bao X C, Chen Q L. Oak forest in Beijing mountainous area. Acta Phytoecologica et Geobotanica Sinica, 1985, 9(2): 101-111. |
| [3] | Wang W, Liu C R, Ma K P, Yu S L. Population structure and dynamics of Quercus liaotungensis in two broad-leaved deciduous forests in Dongling Mountain, northern China. Acta Botanica Sinica, 1999, 41(4): 425-432. |
| [4] | Shi Z M, Liu S R, Cheng R M, Jiang Y X. Interspecific association of plant populations in deciduous broad-leaved forest in Baotianman. Scientia Silvae Sinica, 2001, 37(2): 29-35. |
| [5] | Wu Z Y. Chinese Vegetation. Beijing: Science Press, 1980. |
| [6] | Xie J Y, Chen L Z. Species diversity characteristics of deciduous forests in the warm temperate zone of north China. Acta Ecologica Sinica, 1994, 14(4): 337-344. |
| [7] | Jin Z X. Dominant population structure and trend of deciduous broad-leaved forest in the Tiantai Mountains of Zhejiang. Journal of Zhejiang Forestry College, 2001, 18(3): 245-251. |
| [8] | Shen Z H, Jin Y X, Zhao Z E, Wu J Q, Huang H D. The structure and dynamics of the rare plant communities in subtropical mountain of China. Acta Ecologica Sinica, 2000, 20(5): 800-807. |
| [9] | Yang Y C, Da L J, You W H. Vegetation structure in relation to micro-landform in Tiantong National Forest Park, Zhejiang, China. Acta Ecologica Sinica, 2005, 25(11): 2830-2840. |
| [10] | Shang K K, Chen B, Da L J. Differentiation pattern and topographic interpretation of tertiary relict deciduous forests on Mt. Tianmushan, Zhejiang province, China. Ecology and Environmental Sciences, 2013, 22(2): 213-221. |
| [11] | Zeng S Q, Li J, Li D L, Zhang M, Huang Y J. Study on diameter structure of Choerospondias axillaris secondary forest in southern collective forest area. Journal of Central South University of Forestry & Technology, 2012, 32(4): 1-6. |
| [12] | Chen C J, Chen H F. Planting broad-leaved forest. Beijing: China Forestry Publishing, 2000. |
| [13] | Liu X G, Xie X H, Wen F H, Xie X F. Choerospondias axillaries, the economic forest in south of Jiangxi. Territory & Natural Resources Study, 1995, (2): 64-67. |
| [14] | Li C W, Cui C B, Cai B, Yao Z W. Study progress of Choerospondias axillaries. Pharmaceutical Journal of China People’s Liberation Army, 2008, 24(3): 231-234. |
| [15] | Wei Y. Investigation on the vegetation of Dashanchong forest park in Changsha County. Hunan Forestry Science & Technology, 2009, 36(2): 59-61. |
| [16] | Liu C, Xiang W H, Tian D L, Fang X, Peng C H. Overyielding of fine root biomass as increasing plant species richness in subtropical forests in central southern China. Chinese Journal of Plant Ecology, 2011, 35(5): 539-550. |
| [17] | Qi C J, Yu X L. Seed Plants the Overview of Hunan. Changsha: Hunan Science and Technology Press, 2002: 1-589. |
| [18] | Li B, Yang C, Lin P. Ecology. Beijing: Higher Education Press, 2000. |
| [19] | Li J, Study on structural characteristics and growth model of typical stand types in southern collective forest region. Changsha: Central South University of Forestry and Technology. 2012. |
| [20] | Lei S K, Wang Y J, Wang Y Q, Zhu W L, Yi Q, Feng H M. Photosynthetic physiological characteristics of three typical broad-leaved trees in Jinyun Mountain. Research of Soil and Water Conservation, 2012, 19(2): 189-194, 200-200. |
| [21] | Chen J Z, Zhang S S. A leurites montana, the renewal and repairing wood species in ecological public welfare forests. Subtropical Agriculture Research, 2008, 4(2): 65-67. |
| [22] | Li J, Zhang J. Advances in studies of Litsea coreana levl. var. Journal of Sichuan Agricultural University, 2005, 23(2): 247-252. |
| [23] | Peng S L. South Asian Tropical Forest Community Dynamics. Beijing: Science Press, 1996. |
| [24] | Zhu X W, Zhang K R, Liu K M, Wang S L, Tian S Z, Liu Y D. The quantitative classification of plant communities in Yuelu mountain. Journal of Natural Science of Hunan Normal University, 2009, 32(3): 89-94. |
| [25] | Zhang J, Guo X Y, Da L J, Wang J, Hui X P, Geng Y. Regeneration pattern and limitation of understory woody plants of artificial forest in Shanghai. Journal of East China Normal University: Natural Science, 2013, (2): 11-19. |
| [1] | 中国植被编委会. 中国植被. 北京: 科学出版社, 1980. |
| [2] | 陈灵芝, 鲍显诚, 陈清朗. 北京山区的栎林. 植物生态学与地植物学丛刊, 1985, 9(2): 101- 111. |
| [3] | 王巍, 刘灿然, 马克平, 于顺利. 东灵山两个落叶阔叶林中辽东栎种群结构和动态. 植物学报, 1999, 41(4): 425- 432. |
| [4] | 史作民, 刘世荣, 程瑞梅, 蒋有绪. 宝天曼落叶阔叶林种间联结性研究. 林业科学, 2001, 37(2): 29- 35. |
| [5] | 吴征镒. 中国植被. 北京: 科学出版社, 1980. |
| [6] | 谢晋阳, 陈灵芝. 暖温带落叶阔叶林的物种多样性特征. 生态学报, 1994, 14(4): 337- 344. |
| [7] | 金则新. 浙江天台山落叶阔叶林优势种群结构与动态分析. 浙江林学院学报, 2001, 18(3): 245- 251. |
| [8] | 沈泽昊, 金义兴, 赵子恩, 吴金清, 黄汉东. 亚热带山地森林珍稀植物群落的结构与动态. 生态学报, 2000, 20(5): 800- 807. |
| [9] | 杨永川, 达良俊, 由文辉. 浙江天童国家森林公园微地形与植被结构的关系. 生态学报, 2005, 25(11): 2830-2840. |
| [10] | 商侃侃, 陈波, 达良俊. 浙江天目山孑遗落叶阔叶林群落分异格局及其地形解释. 生态环境学报, 2013, 22(2): 213- 221. |
| [11] | 曾思齐, 李俊, 李东丽, 张敏, 黄炎俊. 南方集体林区南酸枣次生林林分结构研究. 中南林业科技大学学报, 2012, 32(4): 1- 6. |
| [12] | 陈存及, 陈伙法. 阔叶树种栽培. 北京: 中国林业出版社, 2000. |
| [13] | 刘晓庚, 谢兴红, 文芳华, 谢喜发. 赣南山区经济林——南酸枣. 国土与自然资源研究, 1995, (2): 64- 67. |
| [14] | 李长伟, 崔承彬, 蔡兵, 姚志伟. 南酸枣的研究进展. 解放军药学学报, 2008, 24(3): 231- 234. |
| [15] | 魏勇. 长沙县大山冲森林公园植被调查. 湖南林业科技, 2009, 36(2): 59- 61. |
| [16] | 刘聪, 项文化, 田大伦, 方晰, 彭长辉. 中亚热带森林植物多样性增加导致细根生物量“超产”. 植物生态学报, 2011, 35(5): 539- 550. |
| [17] | 祁承经, 喻勋林. 湖南种子植物总览. 长沙: 湖南科学技术出版社, 2002: 1- 589. |
| [18] | 李博, 杨持, 林鹏. 生态学. 北京: 高等教育出版社, 2000. |
| [19] | 李俊. 南方集体林区典型林分类型结构特征及生长模型研究. 长沙: 中南林业科技大学, 2012. |
| [20] | 雷声坤, 王玉杰, 王云琦, 朱蔚利, 易琴, 冯华敏. 缙云山三种典型阔叶树光合生理特性研究. 水土保持研究, 2012, 19(2): 189- 194, 200- 200. |
| [21] | 陈建忠, 张水生. 生态公益林更新和补植树种——千年桐. 亚热带农业研究, 2008, 4(2): 65- 67. |
| [22] | 李俊, 张健. 毛豹皮樟的研究进展. 四川农业大学学报, 2005, 23(2): 247- 252. |
| [23] | 彭少麟. 南亚热带森林群落动态学. 北京: 科学出版社, 1996. |
| [24] | 朱晓文, 张克荣, 刘克明, 汪思龙, 田淑珍, 刘应迪. 岳麓山风景名胜区植物群落数量分类. 湖南师范大学自然科学学报, 2009, 32(3): 89- 94. |
| [25] | 张净, 郭雪艳, 达良俊, 王婕, 惠晓萍, 耿晔. 上海人工林林下木本植物更新格局及其影响因素. 华东师范大学学报: 自然科学版, 2013, (2): 11- 19. |