 2014, Vol. 34
2014, Vol. 34文章信息
- 张淑花, 张雪萍
- ZHANG Shuhua, ZHANG Xueping
- 基于δ15N稳定同位素分析的人工防护林大型土壤动物营养级研究
- Study on the trophic levels of soil macrofauna in artificial protection forests by means of stable nitrogen isotopes
- 生态学报, 2014, 34(11): 2892-2899
- Acta Ecologica Sinica, 2014, 34(11): 2892-2899
- http://dx.doi.org/10.5846/stxb201310132460
-
文章历史
- 收稿日期:2013-10-13
- 网络出版日期:2014-4-3






2. 绥化学院农业与水利工程学院, 绥化 152061
2. College of Agricultural and Hydraulic Engineering, Suihua University, Heilongjiang Suihua 152061, China
关于生态系统的物质循环和能量流动方面的研究一直是生态学者研究的重点,土壤动物作为土壤生物的重要组成部分,在土壤生态系统中发挥着重要的作用。生物之间营养关系是生物群落内各成员之间最为重要的联系方式,是群落赖以生存的基础,也是生态系统能量流动的核心[1, 2]。土壤动物之间的营养关系极其复杂,因其种类多、数量大、个体小以及栖息于地下环境等特点,使得人们关于土壤动物食物链和食物网的认识一直处于模糊状态[3]。传统研究营养关系的方法是通过对消化道内的食物成分进行分析来对食物进行确定,但因为不同的食物类型被消化吸收的速度不同,吸收速度快的食物的作用常常被低估[4, 5],而且对于土壤动物等这些体型较小的动物很难对他们进行胃含物的分析。
稳定同位素是天然存在于生物体内的不具有放射性的一类同位素,早在20世纪70年代末期稳定同位素技术就被引入到生态学研究领域,随着同位素技术应用越来越成熟,其在生态学研究中的重要性也日益突出。近年来,稳定性同位素技术被应用于分析生态系统中的营养关系,成为一种简单又实用,并且准确度较高的一种方法[6],对推动生态系统功能研究具有重要意义。在食物链传递过程中,氮稳定同位素(15N)通常随着营养等级升高而富集,并且每个营养级间具有相对恒定(1.5‰—3.0‰)的富集效应[7, 8],通过对生态系统中不同生物类群δ15N进行测定,能够准确的分析出不同生物的营养级位置,在研究生态系统食物结构及营养关系方面具有强大的作用,尤其是对一些体型较小生物类群的研究更体现出其独有的优势,目前该技术在很多类型生态系统的功能分析及食物网研究中都有一定的应用[9, 10, 11, 12, 13],但该技术的应用还有很多不成熟的方面,如不同生态系统中生物δ15N富集量的差异性、影响生物δ15N值的因素等还没有一致的认识[8, 14]。土壤动物功能研究一直都是土壤动物生态学研究的重要内容之一,传统的研究方法无法对土壤动物之间的营养关系进行精确的认识,稳定同位素的应用使这一问题可以得到解决,但该研究方法在土壤生态系统研究中的应用还处于刚刚开始阶段,虽然如此,其在土壤动物营养关系研究方面的强大的功能和独特的优势已经被很多研究人员所认识,目前国外已有不少相关方面的研究报道[14, 15, 16, 17, 18]。我国土壤动物研究起步较晚,在土壤动物功能方面研究还很薄弱,土壤动物营养关系的研究主要以定性划分功能类群为主[19, 20],定量确定不同土壤动物的营养级及它们之间的营养关系还没有研究,稳定同位素技术在土壤动物研究方面的应用目前还未有报道。
以杨树为主的人工防护林在防风固沙等方面发挥了重要作用[21],但在不同的环境条件下,其群落功能及林下生境的变化对土壤生态系统具有重要的影响,土壤动物作为土壤生态系统的重要组成部分和生态系统健康的重要指示生物,其群落组成与功能变化也是对防护林生态系统变化的一种响应。在先前对土壤动物功能类群研究有一定认识的基础上[19, 20],探索使用稳定氮同位素技术来分析土壤动物的功能关系,选择3处人工防护林作为土壤动物取样地,对其大型土壤动物的δ15N及全氮值进行测定,同时测定了环境中土壤、细根及枯落叶的相应值,在此基础上分析了不同环境条件下土壤动物δ15N的差异性及与其它因素之间的关系,并对不同土壤动物的营养等级进行划分。本文研究旨在探索应用15N稳定同位素技术来划分土壤动物的营养等级,为推动土壤动物功能研究进一步发展,深入分析土壤动物的食物链关系和食物网结构及营养元素迁移转化奠定一定的基础。
1 材料与方法 1.1 研究区域概况研究区域位于黑龙江省哈尔滨市的道外区和阿城区境内,属于东北松嫩平原和东部山地交接地带,地貌类型以平原和低山丘陵为主;气候属于中温带大陆性半湿润季风气候,年平均气温为3.4 ℃,多年平均降水量为500—600 mm,降水主要集中在6—8月,约占全年降水量的65%,但不同年份的降水变率较大;土壤类型主要以黑土、草甸土等为主,0—5 cm层土壤有机质含量为5%—10%,为肥力水平较高的土壤。在研究区域内选择3处典型的人工防护林作为采样地,具体试验样地的环境特征见表1。
| 样地 Sample plots | 地理位置 Geographic position | 海拔高度 Height above sea level/m | 防护林类型 Protection forest type | 防护林树高 Height of tree/m | 造林年份 Forestation year | 凋落物厚度 Forest litter thickness | 土壤pH值 pH value of soil | 表土层有机质/% Soil organic matter/% | 表土层含水量 Soil moisture content/% |
| 万宝镇 | 45°51′N,126°25′E | 118 | 杨树 | 25 | 2002 | 2 cm | 7.03 | 6.93 | 15.27 |
| 杨树乡 | 45°28′N,126°50′E | 180 | 杨树 | 16 | 2005 | 3—4 cm | 7.22 | 5.80 | 8.83 |
| 平山镇 | 45°19′N,127°23′E | 240 | 杨树 | 20 | 2003 | 5—6 cm | 7.19 | 5.69 | 13.67 |
于2010年9月在所选样地内选择有代表性的地点设置样方8处,每个样方面积约为25 cm×50 cm,对样方内土壤动物用手捡法进行分离,所分离出的土壤动物马上放入75%的酒精内进行杀死固定,带回室内进行分类鉴定。在3个样地获得的土壤动物中,选择共有的12个动物类群,对不同类群土壤动物逐一进行处理,蚯蚓、大蚊幼虫及金龟甲幼虫因其体内含有较多未消化物质,对这几类动物进行解剖,将肠道内未消化物剔除,其它动物类群未进行解剖处理,处理完的样品在60 ℃下烘干24 h,并分别放入密封袋内使其保持干燥。植物细根、枯落叶用水将杂质清洗干净后,40 ℃条件下烘干,同样放入密封袋内;风干后的土壤样品用100目土壤筛进行处理后放入95 ℃恒温箱内烘干,置于密封袋内待进行同位素及全氮分析。
1.3 稳定同位素及全氮分析试验样品的稳定同位素分析是在中国林科院稳定同位素比率质谱实验室进行,采用的仪器为美国Thermo Fisher Scientific公司的MAT253同位素比率质谱仪进行测定,氮稳定同位素比值以国际通用的δ值形式表达,测定原理为将样品在元素分析仪中高温燃烧后生成N2,质谱仪通过检测N2的15N和14N比率,并与国际标准物(大气N2)对比后计算出样品的δ15N比率值,δ15N计算的公式为:
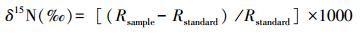
营养等级确定的公式为:
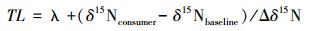
环境条件的差异使不同生态系统具有不同的δ15N营养等级富集度,有研究认为δ15N富集度的平均数为3.4‰[22],但学者对不同地区、不同类群的动物研究得出不同的结果[9, 10, 11, 12, 13],植食性、肉食性和杂食性生物对δ15N的富集量没有明显的区别,但腐食性生物对δ15N的富集效应明显偏低[9, 23];Tiuno A.V.对不同地区不同类群土壤动物随食物链的富集状况进行了总结,结果显示第一级消费者的δ15N比其食物增加0—5‰,捕食性生物相对于所捕食食物来说δ15N富集量约为2.4‰—7.6‰[14]。
在本研究的3个样地中,土壤动物δ15N值最低的是大蚊幼虫,而大多数大蚊幼虫是取食植物根系或是腐败植物,因此本文将其定为初级消费者,3个样地大蚊幼虫与枯落叶和植物细根之间的平均差值分别为1.86‰和0.48‰,结合前人对δ15N在相邻营养级的富集量的研究[8, 14],本文选择2.3‰作为富集量进行分析。将数据带入公式,即得到下式:
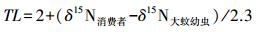
本文相关系数采用Pearson相关分析,使用双尾检验(2-tailed)其相关显著性,P>0.05表明不具有相关性,P<0.05表明显著相关,P<0.01表明极显著相关。不同样地之间的差异性以单因素方差分析(F检验)进行统计,P>0.05说明无显著差异,P<0.05说明差异显著,P<0.01说明差异极显著。文中数据处理和统计分析主要使用SPSS和Excel软件进行。
2 结果与分析 2.1 结果 2.1.1 δ15N测定结果利用同位素质谱仪对3处防护林样地的土壤、细根、枯落叶及12种土壤动物进行了氮稳定同位素测定,将其与标准值相比较得出δ15N结果见图 1。从图 1可以看出,不论是在哪个样地,土壤动物δ15N值的跨度范围均较大,万宝镇土壤动物δ15N值范围在2.00‰—6.40‰之间,δ15N值最小的是大蚊幼虫(Tipulidae larvae),最大的为线蚓;杨树乡δ15N范围在-0.50‰—4.10‰之间,最小值是蚯蚓,最大值是蜘蛛;平山镇δ15N范围在4.20‰—9.96‰之间,最小值为大蚊幼虫,最大值为隐翅甲成虫,不同土壤动物δ15N值在3个样地变化趋势有一定的差异,但整体上表现出一定的相似性,叩甲科(Elateridae)幼虫及成虫、线蚓、蜈蚣(Scolopendromorpha)、隐翅甲等动物的δ15N值较高,而大蚊幼虫、金龟甲幼虫(Scarabaeidae)、蚯蚓等δ15N值较低,这是由于不同类群的土壤动物在土壤生态系统中具有不同的食性特征,处于不同的营养级位置。3个样地中土壤动物的δ15N平均值相差较大,万宝镇土壤动物δ15N的平均值为5.34‰,杨树乡为1.88‰,平山镇为8.45‰,平均值相差值最大达6.57‰。

|
| 图 1 不同取样地土壤动物及环境的δ15N值 Fig. 1 The δ15N values of soil fauna and environment in different samples |
对3个样地的土壤、枯落叶和细根的δ15N值进行分析,发现在3个样地中土壤的δ15N均高出枯落叶和细根的值,δ15N值大小变化顺序表现为枯落叶<细根<土壤,这与Pollierer M. M.等人对瑞士森林生态系统的研究结果一致[3]。
2.1.2 全氮测定结果对不同样地的土壤动物和环境中全氮进行测定,结果见图 2,从图 2可以看出,土壤动物在不同样地全氮值差异不明显(F=0.190,P=0.828),土壤、枯落叶和细根的全氮值也有相同的表现,对3个样地的全氮值进行相关性分析显示,万宝镇与杨树乡之间相关系数r=0.920(P<0.01),与平山镇的相关系数为r=0.963(P<0.01),杨树乡与平山镇的相关系数r=0.974(P<0.01),说明全氮含量在不同样地之间呈现明显的正相关性。

|
| 图 2 不同取样地土壤动物及环境的全氮值 Fig. 2 The total nitrogen of soil fauna and environment in different samples |
分别对3个样地的土壤动物δ15N值与土壤、枯落叶及细根δ15N值进行相关分析,结果显示不论是哪种土壤动物,其δ15N值与土壤、枯落叶及细根之间都具有明显的相关性,相关系数R值均在0.6以上(P<0.05或P<0.01);对不同样地中土壤动物δ15N值进行差异分析显示,不同样地之间土壤动物δ15N值差异显著(F=38.067,P<0.001),这说明样地环境的δ15N值对土壤动物δ15N具有一定的影响。
对不同样地的土壤动物和环境中δ15N值与他们相应的全氮值进行相关性分析,结果显示万宝镇δ15N值与全氮值的相关系数r=0.571(P=0.026),杨树乡和平山镇δ15N值与全氮值的相关系数分别为0.295(P=0.285)和0.404(P=0.135),这说明分析样品中δ15N值与其全氮含量无明显的相关性,这与Chahartaghi M.等人对于德国黑森州(Hesse-Darmstadt)和下萨克斯州(Lower Saxony)的3处不同山毛榉(栎-山毛榉)样地中跳虫的δ15N和全氮关系的分析结果相同[16]。
对3个样地的土壤动物δ15N值进行相关性分析显示万宝镇和平山镇及杨树乡之间相关性明显,相关系数分别为r=0.819(P=0.001)和r=0.771(P=0.003),平山镇和万宝镇之间的相关系数r=0.466(P=0.127),由此可以说明,虽然土壤动物δ15N值在不同样地之间差异较大,但它们之间的联系在不同样地中具有一些相同表现,不同样地土壤动物δ15N值的相关系数较高,这使得通过分析不同土壤动物类群δ15N值来确定其营养级成为可能。
2.2.2 土壤动物营养级分析本研究对12类大型土壤动物的δ15N进行分析,它们的营养关系有直接联系,但更多情况是间接的联系,基于此,仅对它们进行营养级的划分而未进行食物网关系的确定。采用前述方法对12种大型土壤动物的营养级进行划分,结果见图 3。

|
| 图 3 不同类型土壤动物营养级 Fig. 3 The nutrition levels of different types of soil fauna |
对图 3分析可以看出,参与试验的12类大型土壤动物营养级差异较大,大多数类群是处于第2—4营养级之间,也有超过第4营养级,其中处于较低营养级的类群主要有大蚊幼虫、金龟甲幼虫、蚯蚓等,处于较高营养级的动物主要有蜈蚣、隐翅甲成虫、叩甲成虫、线蚓等类群。同一类动物在不同样地营养级划分上有一定的差异,有的甚至相差1个以上的营养级,原因是由于每类土壤动物在不同样地存在取食方面的差异性,此外分类不够细化也是导致其营养级差异较大的原因。
3 讨论同一类群的不同物种在食性方面可能有很大的差别,导致它们之间δ15N值也可能存在很大的差别,因此,在运用δ15N分析土壤动物营养级的研究中一般对于生物分类要求较高,一般要求划分到种的分类水平[14]。但很多同属于一个类群的动物在食性方面具有很大的相似性,因此通过氮稳定同位素分析方法能够大致确定不同类群土壤动物的营养级位置,但要想准确的分析生物之间的营养关系,对生物分类方面就有着较严格的要求[14, 18]。本文通过对几类大型土壤动物及其生存环境的土壤、细根和枯落叶的δ15N及全氮含量进行分析,能够对几类土壤动物的营养级位置初步确定,并对影响土壤动物δ15N的因素进行分析。
影响土壤有机氮同位素组成的因素包括早期成岩作用和其它环境因子以及有机氮的来源、外源有机氮的输入等方面[24, 25],土壤动物生存地δ15N值直接影响其食物的δ15N值的大小[16, 24],这导致不同样地土壤动物δ15N值差异较大,本文研究结果显示3个研究样地土壤动物δ15N差异显著(F=38.067,P<0.001)。土壤动物δ15N值与土壤、植物细根和枯落叶的δ15N值之间具有明显的相关性,与土壤动物体内的全氮含量间不具有相关性[16],这一点在本研究中也得到了证实。3个样地的土壤动物δ15N值具有明显的相关性,这为通过分析不同土壤动物类群δ15N值来确定其营养级提供可能条件。土壤是自然界中物种丰富度最高的生态系统之一,因其生物种类和数量多、个体小等原因,导致到目前为止人们对于土壤生态系统中生物之间关系的研究一直处于模糊状态[3],这严重阻碍了土壤生态系统功能研究的进展,随着现代先进技术手段在生态学中的应用,尤其是稳定同位素技术的应用,使得探索土壤生物之间的功能关系有了可能[6, 25]。土壤生态系统作为地下生态系统,其动物的营养来源与地表生态系统有明显的差异,在地表陆地上第一营养级主要是植物,在水生生态系统中,第一营养级主要是浮游生物[4],但在土壤生态系统中第一营养级可能是新鲜的植物,可能是腐败的枯落叶,也可能是真菌等微生物,而且在自然界中一种生物的食物源往往有多种类型[26],同一土壤动物在不同时空条件下可能有着不同的取食对象,有时不同的土壤动物还会取食相同的食物,这使得土壤动物食物网结构非常复杂[27]。根据δ15N值在食物链传递过程中具有富集效应的理论能够划分土壤动物的营养级,但不同类群生物对δ15N富集程度差异较大,软体动物富集量为1.2‰,甲壳类动物约为2.0‰,昆虫约为2.7‰,而脊椎动物约为2.4‰—3.0‰,捕食性动物、植食性动物和杂食性动物的δ15N富集量差异不大,分别为2.69‰、2.98‰和2.56‰,而腐食性动物的富集量明显偏低(0.53‰)[14, 28, 29],即便是同一种类群,在不同的研究区域也存在一定差异性,如同样对跳虫的富集量研究存在很大的差异,Ruess L.等研究的富集量为-2.0‰—3.5‰[30],而Haubert D.等研究的富集量为2.4‰—6.3‰[31],本文以大蚊幼虫作为初级消费者,结合前人对δ15N富集量的研究及本试验结果,选择2.3‰作为土壤动物δ15N富集量,比大蚊幼虫与枯落叶和细根之间的δ15N差值稍大,这使土壤动物营养级的位置可能偏低。
稳定同位素技术的发展为生态学者进一步深入研究生态系统特征与过程提供了条件,应用δ15N分析土壤动物营养关系方面的研究还有很多问题需要探讨,仅依靠δ15N很难准确的分析出不同生物之间的营养关系,很多研究将δ15N与δ13C结合进行生物营养关系的研究[3, 9, 13, 15],由于自然条件下生态系统食物网关系的复杂性,仅依靠碳、氮稳定同位素分析也很难完成对食物网结构的全面认识,多种同位素协同分析(如δ13C、δ15N、δ34S、δD等)可能会更有助于认清生态系统的食物网关系,但这方面的研究还处于探索阶段[14, 32, 33, 34]。本文应用15N稳定同位素技术分析不同类型土壤动物营养级及δ15N的影响因素,是同位素技术在土壤动物营养关系研究方面应用的初步探讨,因调查的土壤动物种类有限,只涉及部分大型土壤动物而没有包括中型及小型类群,分类研究工作进行的也不够深入,土壤动物的食物源只分析了土壤、细根和枯落叶,其它可能的食物源(如细菌、真菌、藻类等)没有进行分析,这些因素导致本研究未能对土壤动物的食物网关系进行分析。
| [1] | Fu S L. A reviw and perspective on soil biodiversity research. Biodiversity Science, 2007,15(2):109-115. |
| [2] | Yin X Q, Song B, Dong W H, Xin W D, Wang Y Q. A review on the eco-geography of soil fauna in China. Journal of Geographical Sciences, 2010,20(3):333-346. |
| [3] | Pollierer M M, Langel R, Scheu S, Maraun M. Compartmentalization of the soil animal food web as indicated by dual analysis of stable isotope ratios (15N/14N and 13C/12C). Soil Biology and Biochemistry, 2009, 41(6): 1221-1226. |
| [4] | Yang J M. A study on food and trophic levels of Bohai Sea invertebrates. Modern Fisheries Information, 2001,16(9):8-16. |
| [5] | Yan Y R, Lu H S, Jin X S. Marine fish feeding ecology and food web: Progress and perspectives. Journal of Fisheries of China, 2011,35(1):145-153. |
| [6] | Yi X F, Zhang X A. Application of stable isotopic approach in ecology: a review. Chinese Journal of Ecology,2005,24(3):306-314. |
| [7] | Hood-Nowotny R, Knols B G J. Stable isotope methods in biological and ecological studies of arthropods. Entomologia Experimentalis et Applicata, 2007, 124(1): 3-16. |
| [8] | McCutchan J H, Lewis W M, Kendall C, McGrath C C. Variation in trophic shift for stable isotope ratios of carbon, nitrogen, and sulfur. Oikos, 2003, 102(2): 378-390. |
| [9] | Li D H, Su X M. The studies on the food web structures and trophic relationships in Guangxi Dongfang Cave by means of stable carbon and nitrogen isotopes. Acta Ecologica Sinica, 2012, 32(11): 3497-3504. |
| [10] | Ke Z X, Hung L M, Xu J, Ma Y E. The stable nitrogen isotope of size-fractioned plankton and its relationship with biomass during winter in Daya Bay. Acta Ecologica Sinica, 2012,32(22): 7102-7108. |
| [11] | Darby B J, Neher D A. Stable isotope composition of microfauna supports the occurrence of biologically fixed nitrogen from cyanobacteria in desert soil food webs. Journal of Arid Environments, 2012, 85: 76-78. |
| [12] | Poste A E, Muir D C G, Mbabazi D, Hechy R E. Food web structure and mercury trophodynamics in two contrasting embayments in northern Lake Victoria. Journal of Great Lakes Research, 2012, 38(4): 699-707. |
| [13] | Klamer B, Maraun M, Scheu S. Trophic diversity and niche partitioning in a species rich predator guild-Natural variations in stable isotope ratios (13C/12C, 15N/14N) of mesostigmatid mites (Acari, Mesostigmata) from Central European beech forests. Soil Biology and Biochemistry, 2013, 57: 327-333. |
| [14] | Tiuno A V. Stable isotopes of carbon and nitrogen in soil ecological studies. Biology Bulletin, 2007, 34(4): 395-407. |
| [15] | Heiner B, Drapela T, Frank T, Zaller J G. Stable isotope 15N and 13C labelling of different functional groups of earthworms and their casts: A tool for studying trophic links. Pedobiologia, 2011, 54(3): 169-175. |
| [16] | Chahartaghi M, Langel R, Scheu S, Ruess L. Feeding guilds in Collembola based on nitrogen stable isotope ratios. Soil Biology and Biochemistry, 2005, 37(9): 1718-1725. |
| [17] | Caner L, Zeller B, Dambrine E Ponge J F, Chauvat M, Llanque C. Origin of the nitrogen assimilated by soil fauna living in decomposing beech litter. Soil Biology & Biochemistry, 2004, 36(11): 1861-1872. |
| [18] | Scheu S, Falca M. The soil food web of two beech forests (Fagus sylvatica) of contrasting humus type: stable isotope analysis of a macro-and a mesofauna-dominated community. Oecologia, 2000,123(2):285-296. |
| [19] | Zhang X P, Hou W L, Chen P. Soil animal guilds and their ecological distribution in the northeast of China. Chinese Journal of Applied & Environmental Biology, 2001,7(4):370-374. |
| [20] | Zhang S H, Zhang X P. Guilds of soil macro-animals in burned forest areas in Daxing'an Mountains. Journal of Northeast Forestry University, 2011,39(8):70-72. |
| [21] | Du H Q,Han Z W,Yan C Z,Deng X H, Song X, Liao J. Effects of wind prevention by the northwest shelterbelt. Bulletin of Soil and Water Conservation, 2010,30(1):117-120. |
| [22] | Vander Zanden M J, Rasmussen J B. Variation in δ15N and δ13C trophic fractionation: implications for aquatic food web studies. Limnology and Oceanography, 2001, 46(8): 2061-2066. |
| [23] | Vanderklift M A, Ponsard S. Sources of variation in consumer-diet δ15N enrichment: a meta-analysis. Oecologia, 2003,136(2): 169-182. |
| [24] | Briones M J I, Bol R, Sleep D, Allen D, Sampedro L. Spatio-temporal variation of stable isotope ratios in earthworms under grassland and maize cropping systems. Soil Biology & Biochemistry, 2001, 33(12/13):1673-1682. |
| [25] | Lin G H. Stable isotope ecology: a new branch of ecology resulted from technology advances. Chinese Journal of Plant Ecology, 2010, 34 (2): 119-122. |
| [26] | Phillips D L, Gregg J W, Source partitioning using stable isotopes: coping with too many sources. Oecologia, 2003,136(2): 261-269. |
| [27] | Liu C H, Liu S P, Wang X J, Chen Z L, Lian Z M. The soil fauna guilds and its ecological regulation of pests at jujube forest in northern region of Shanxi. Chinese Journal of Soil Science, 2011,42(2):316-319. |
| [28] | Halaj J, Peck R W, Niwa C G. Trophic structure of a macroarthropod litter food web in managed coniferous forest stands: a stable isotope analysis with δ15N and δ13C. Pedobiologia, 2005, 49(2):109-118. |
| [29] | Seeber J, Seeber G U H, Kössler W, Langel R, Scheu S, Meyer E. Abundance and trophic structure of macro-decomposers on alpine pastureland(Central Alps, Tyrol): effects of abandonment of pasturing. Pedobiologia, 2005, 49(3): 221-228. |
| [30] | Ruess L, Häggblom M M, Langel R, Scheu S. Nitrogen isotope ratios and fatty acid composition as indicators of animal diets in belowground systems. Oecologia, 2004, 139(3): 336-346. |
| [31] | Haubert D, Langel R, Scheu S, Ruess L. Effects of food quality, starvation and life stage on stable isotope fractionation in Collembola. Pedobiologia, 2005, 49(3): 229-237. |
| [32] | Polierer M M, Langel R, Körner C, Maraun M, Scheu S. The underestimated importance of belowground carbon input for forest soil animal food webs. Ecology Letters, 2007,10(8):729-736. |
| [33] | Arneson L S, MacAvoy S E. Carbon, nitrogen, and sulfur diet-tissue discrimination in mouse tissues. Canadian Journal of Zoology, 2005,83(7):989-995. |
| [34] | Birchall J, O'Connell T C, Heaton T H E, Hedges R E M. Hydrogen isotope ratios in animal body protein reflect trophic level. Journal of Animal Ecology, 2005,74(5):877-881. |
| [1] | 傅声雷. 土壤生物多样性的研究概况与发展趋势. 生物多样性,2007, 15(2): 109-115. [4] 杨纪明. 渤海无脊椎动物的食性和营养级研究. 现代渔业信息, 2001, 16(9): 8-16. |
| [5] | 颜云榕, 卢伙胜, 金显仕. 海洋鱼类摄食生态与食物网研究进展. 水产学报, 2011, 35(1): 145-153. |
| [6] | 易现峰, 张晓爱. 稳定性同位素技术在生态学上的应用. 生态学杂志, 2005, 24(3): 306-314. |
| [9] | 黎道洪, 苏晓梅. 应用稳定同位素研究广西东方洞食物网结构和营养级关系. 生态学报, 2012, 32(11): 3404-3497. |
| [10] | 柯志新, 黄良民, 徐军,马艳娥. 大亚湾冬季不同粒级浮游生物的氮稳定同位素特征及其与生物量的关系. 生态学报, 2012, 32(22): 7102-7108. |
| [19] | 张雪萍, 侯威岭, 陈鹏. 东北森林土壤动物同功能种团及其生态分布. 应用与环境生物学报, 2001,7(4): 370-374. |
| [20] | 张淑花, 张雪萍. 大兴安岭森林火烧迹地大型土壤动物功能类群. 东北林业大学学报, 2011,39(8): 70-72. |
| [21] | 杜鹤强, 韩致文, 颜长珍, 邓晓红, 宋翔, 廖杰. 西北防护林防风效应研究. 水土保持通报, 2010,30(1):117-120. |
| [25] | 林光辉. 稳定同位素生态学: 先进技术推动的生态学新分支. 植物生态学报, 2010,34(2):119-122. |
| [27] | 刘长海, 刘世鹏, 王晓涧, 陈宗礼, 廉振民. 陕北枣林土壤动物功能类群及其害虫生态调控. 土壤通报,2011,42(2):316-319. |