 2014, Vol. 34
2014, Vol. 34文章信息
- 石洪华, 王晓丽, 郑伟, 王嫒
- SHI Honghua, WANG Xiaoli, ZHENG Wei, WANG Ai
- 海洋生态系统固碳能力估算方法研究进展
- Review of carbon sequestration assessment method in the marine ecosystem
- 生态学报, 2014, 34(1): 12-22
- Acta Ecologica Sinica, 2014, 34(1): 12-22
- http://dx.doi.org/10.5846/stxb201304280855
-
文章历史
- 收稿日期:2013-4-28
- 修订日期:2013-10-23






2. 天津理工大学环境科学与安全工程学院, 天津 300384
2. College of Environmental Science and Safety Engineering, Tianjin University of Technology, Tianjin 300384, China
CO2对全球气温升高的贡献高达70%,居各种温室气体之首[1]。从生物地球化学的角度看,海洋在全球碳循环中发挥着重要作用[2]。海洋占地球总面积的71%,其CO2贮存量是大气的50倍,是大气CO2的调节器。各种模式和实测结果表明,全球海洋对CO2的净吸收能力为1.5—2.0PgC/a,约占人为释放CO2年总量的25%—50%[3, 4, 5, 6, 7]。对海洋碳循环及其固碳能力的评估方法的研究已成为当今海洋学的重要课题。
海洋中的碳主要以碳酸盐离子的形式存在,如溶解无机碳(DIC)、溶解有机碳(DOC)、颗粒有机碳(POC)以及生物有机碳(BOC)[8]。海洋碳循环中最重要的两个过程是物理泵和生物泵[9]。物理泵指发生在海-气界面的CO2气体交换过程和将CO2从海洋表面向深海输送的物理过程,生物泵指浮游生物通过光合作用吸收碳并向深海和海底沉积输送的过程。因而,海洋碳循环的碳通量的估算过程如图1所示[10]。

海-气界面的CO2气体交换,是海洋碳循环中与人类影响密切相关的重要一环,且直接影响大气CO2的含量[11]。通过气体交换从大气进入海洋的CO2的多少主要取决于风速和海-气界面两侧的CO2分压差,同时,由于海水对CO2的溶解度与温度有关,因此海水对CO2的吸收量也是温度的函数[7, 8, 12]。海-气界面CO2交换通量代表海洋吸收或放出CO2的能力。准确估算海-气界面CO2交换通量对深入理解海洋碳循环及预测大气CO2变化具有重要意义。
CO2进入海水后,在真光层内通过浮游生物的光合作用转化成有机碳,其中大部分有机碳停留在上层海洋中通过食物链进行循环,小部分以POC沉降颗粒物的形式从真光层输出而进入海洋深层水体[5, 6]。这部分通过生物泵向深海输送的碳,由于其与大气隔绝,可在百年乃至更长的时间尺度上影响大气CO2含量。而真光层内浮游生物的初级生产力既能影响海-气界面CO2交换通量,也能影响真光层内POC输出通量。因此,浮游生物的初级生产力估算、真光层的POC输出通量也是海洋碳迁移研究的重点,POC输出通量与初级生产力的比值可用于衡量生物泵的运转效率[9]。从真光层输送到深海的有机碳中,一部分被微生物分解还原为CO2,只有很小一部分被埋藏在海底沉积物中长期封存。在一定时间尺度内,海洋“生物泵”引起的沉积有机碳埋藏可以认为是海洋碳元素的最终归宿[13],因而海洋有机碳的沉积通量可认为是海洋碳汇作用的最终效应。
海-气界面CO2气体交换通量、浮游生物的初级生产力、真光层POC输出通量以及有机碳沉积通量是海洋碳循环过程中的重要评估参数,准确估算它们的大小及其比例关系,能有效的说明海洋的生物泵运转效率和海洋在不同时间尺度上的碳汇效应。本文重点综述了海洋海-气界面CO2气体交换通量、浮游生物的初级生产力、真光层POC输出通量以及有机碳沉积通量的估算方法,分析了海洋生态系统固碳估算方法的应用中存在的问题,并展望了海洋生态系统固碳估算方法的发展方向。
1 海-气界面CO2气体交换通量估算方法研究大气中的CO2进入海洋后,在海洋-大气界面通常存在一个CO2浓度梯度,在大气和洋流的综合作用下,界面上进行着大量CO2交换。CO2从大气中溶入海水的过程称为“溶解度泵”,其固碳能力估算常采用测算海-气界面CO2通量的方法而获得[14]。海-气界面CO2的源和汇主要是由表层海水CO2分压(PCO2)的分布变化引起的,间接地受到海水温度、生物活动和海水运动等因素的影响[15]。
海-气界面CO2气体交换通量[16]指的是单位时间单位面积上CO2在大气和海洋界面的净交换量。该气体交换通量是评估海洋在全球变化中作用的前提和基础。估算海-气界面CO2交换通量方法一般分为两类,一类为包括放射性同位素14C示踪法[17]、碳的稳定同位素比例法[18]、通过测量大气O2的镜像法[19]等基于物质守恒原理在全球尺度上估算海-气界面CO2交换通量的方法;另一类分别测量海水和海水表层大气中的CO2分压,结合CO2海-气交换速率来实测-海气界面CO2交换通量。表层海水CO2分压的测量手段包括船载走航测定的水气平衡的非色散红外法、浮标原位时间序列观测的化学传感器法及大时间空间尺度观测的遥感法[20, 21, 22, 23, 24, 25, 26, 27, 28, 29]。测量不同海域的海水和海水表层大气中的CO2分压需要建立海-气界面CO2通量的立体观测平台,该观测平台包括岸基、船基、航空、卫星和浮标等系统,主要技术包括走航大气和海水观测技术、浮标海-气界面CO2通量观测技术、极区海洋-大气界面CO2通量的观测技术和遥感海洋-大气界面CO2通量观测和评估技术,海-气界面CO2通量观测技术方法比较如表 1所示。
|
海-气CO2通量
观测方法 Observation method of sea-air CO2 flux | 工具
Equipment | 系统组成
Composition | 测定主要参数
Main parameters | 空间尺度
Spatial scale | 使用范围
Application scope |
|
走航
Cruise[20-21,29] | 科研调查船、志愿船 | 水-汽平衡器
物化参数传感器 气象数字观测系统 卫星定位系统 数据采集与控制 | 表层海水-气界面CO2分压
温度、盐度、溶解氧和叶绿素等传感器 气压、风速、风向等气象参数 连续走航观测的实时定位测量 控制自动观测、测量数据采集和处理 | 海-气界面小
尺度 | 某一海域 |
|
浮标
Float[26, 28] | 锚系、漂流浮标
| 水-汽平衡器
物化参数传感器 气象数字观测系统 卫星定位系统 数据采集与控制 太阳能电池 | 表层海水-气界面CO2分压
温度、盐度、溶解氧和叶绿素等传感器 气压、风速、风向等气象参数 连续走航观测的实时定位测量 控制自动观测、测量数据采集和处理 浮标等支撑能源体系 | 海-气界面小
尺度 | 某一海域 |
|
遥感Remote
sensing[22, 24, 25, 27] | 卫星 | 气象数字观测系统
卫星定位系统 数据采集和分析 能源电池 | 海面风场和海面高度
观测的实时定位测量 叶绿素浓度、初级生产力、海表温度等支撑能源体系 | 大尺度 | 某一海域
或全球 |
|
走航-浮标-遥感
Cruise-Float-Remote Sensing[23] | 科研调查船、锚系和浮标、卫星 | 水-汽平衡器
物化参数传感器 气象数字观测系统 卫星定位系统 数据采集与控制 能源电池 | 表层海水-气界面CO2分压
温度、盐度、溶解氧和叶绿素等传感器 气压、风速、风向等气象参数 连续走航观测的实时定位测量 控制自动观测、测量数据采集和处理 浮标等支撑能源体系 | 空间尺度 | 极区或
全球 |
采用海-气界面的CO2分压差法估算海-气界面CO2交换通量时,大气和海水的CO2分压都有相对成熟可靠的方法,关键在于气体交换系数的确定,即CO2在海-气界面的传输速率k,这涉及到CO2在海气界面迁移交换这个非常复杂的动力学过程,如近表层水温周日变化、盐度变化、碎浪作用、气泡作用、上升流变动、生物活动、表面温度效应、海表风速、大气边界层性质等都对其有重要影响,而且CO2在海-气界面迁移交换的各种控制机制和过程有显著的时空变化[20, 21, 22, 23, 24, 25, 26, 27, 28, 29]。目前的研究大多假定k主要为风速的函数,但要准确测定某一区域气体交换系数的难度很大,因此,现有研究中多数CO2海-气交换通量估算都是直接引用经典文献给出的风速函数关系[30, 31]。可见,k值一方面缺乏足够精确的现场实测数据,另一方面,不同研究者之间的结果差异巨大,尤其是在高风速区间[20],因为在高风速条件下现场环境恶劣,难以开展实验,而且高风速持续时间一般很短,满足不了开展非直接通量测量方法的需要。为了准确估算全球不同海域的海-气界面CO2交换通量,减少实地测量的局限性和模型预测的不确定性,根据已有记载的不同海域的气候气象资料,利用数学模型模拟和实地测量值不断修正相结合的方法,建立不同海域不同季节的气体交换系数k的动态数据库,以提高海-气界面CO2交换通量估算的准确性。
2 浮游生物的初级生产力估算方法研究海洋的“溶解度泵”只是实现了CO2从大气碳库向海洋碳库的迁移,存在很强的时空异质性。进入海洋的CO2被浮游植物和光合细菌通过光合作用固定转变为有机碳从而进入海洋生态系统,碳在海洋生态系统食物网中经过层层摄食最终以生物碎屑的形式输送到海底,从而实现了碳的封存,封存的碳在几万甚至上百万年时间内不会再进入地球化学循环,这一过程被称为生物泵[32]。生物泵是海洋碳循环中最复杂的,浮游植物和好氧光合细菌通过光合作用固定无机碳,每年大约有45Gt的碳被固定转化为有机碳[33, 34]。固定的碳被浮游动物所摄食成为次级生产力,然后部分被更高营养级生物所摄食,部分通过呼吸和死亡分解再次变成无机碳返回环境,部分被垂直输送到海底,其生产力则占海洋初级生产力的95%以上,其中每年有35Gt有机碳通过生物异养呼吸的途径变成DIC,这部分碳占海洋表面光合作用所固定碳的80%左右[35, 36, 37]。真光层异养细菌是这个过程的主要贡献者,据估计,大约50%—90%的呼吸作用是由异养细菌来完成的[38]。甚至在某些海区,细菌的呼吸作用要强于该地区的初级生产力[39]。未被呼吸作用氧化的有机碳以生物碎屑和排泄物(POC)以及DOC的形式向弱光层、深海无光层输送,每年大约有10Gt有机碳最终进入深海[35],但其中绝大部分经过再矿化再次成为DIC,最终能够进入洋底沉积物的不足5%[40]。
海洋生态系统的碳循环过程主要是通过海洋生物泵完成,而浮游生物的初级生产力是这一过程的起始环节和关键部分[9]。浮游生物固碳强度与潜力可用初级生产力来表征[41]。叶绿素是浮游生物进行光合作用的主要色素,也是海洋中主要初级生产者(浮游生物)现存量的一个良好指标。利用海洋叶绿素浓度测算海洋初级生产力的方法可分为两种模式,即经验统计模型和生态学数理模型。在一定的环境条件下,叶绿素浓度和初级生产力是对应的,存在一定的统计关系。一些研究者在分析海洋叶绿素和初级生产力之间关系的问题时,建立了一系列的经验统计模型,大都为简单的线性关系[42]。经验统计模型的主要局限性是一般只对同一海域适用,精度不高,且随着时间的推移,各参数间的相关性会发生变化。因此,近年来已经很少使用。
从20世纪中期,Ryther和Yentsch开始利用海水中的叶绿素含量建立生态学数理模型来估算海洋浮游植物的初级生产力,为大面积的海洋调查带来了方便[43]。该模型中饱和光条件下浮游植物的光合作用速率是叶绿素浓度的函数,即:
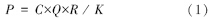
式中,P为浮游植物光合作用速率(mgC m-3 h-1);C为叶绿素浓度(mg/m3);Q为同化系数,是单位质量叶绿素在单位时间内同化的碳量;R为决定于海面光强的相对光合作用率;K为海水消光系数(m-1)。Ryther和Yentsch[42]的研究指出,在上述模式中,标志海洋浮游植物光合作用能力大小的重要参数“同化系数”受各理化因子的影响而具有可变性,这就导致了叶绿素浓度与初级生产力之间的关系不是恒定的。因此,在应用中必须正确地测定调查水域的同化系数。Cadée和Hegeman[44]改进了初级生产力计算模型,其计算公式为:
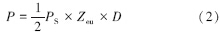
式中,PS为潜在生产力(mgC m-3 h-1),由表层叶绿素浓度和同化系数得到;Zeu为真光层深度(m);D为日照时数(h/d)[45]。潜在生产力PS是表层海水中的叶绿素浓度和同化系数的函数,即为:

式中,Chla为表层叶绿素a的含量(mg/m3),Q为同化系数(mg mg-1 h-1)。同化系数(Q) 是指单位叶绿素a在单位时间内合成的有机碳量,是用来表征浮游生物光合作用强度的量值。浮游生物的同化系数在不同的海域、不同季节差异较大,影响因素除了与不同浮游生物的适应性有关外,还与环境营养盐、光照条件和温度等因素有关,同化系数Q值一般也利用经验关系式估算[46, 47, 48, 49]。
生态学数理模型通过海洋环境因子及海洋叶绿素浓度来估算海洋初级生产力,这种算法模式考虑了光照、水温、营养盐等对海洋初级生产力的直接或间接影响,同时考虑了叶绿素浓度、光照等在垂直剖面上的差异,因而估算得到的海洋初级生产力精度要比经验算法高,且有较强的生物学意义[46, 47, 48]。目前国内海洋浮游植物固碳能力的估算方法一般都采用生态学数理模型[50, 51, 52]。
随着空间探测技术的进步,卫星技术的发展十分迅速。高空间分辨率、高时间分辨率和高光谱分辨率的卫星不断涌现。卫星遥感具有及时、准确、动态和大面积覆盖的特点,因而已逐渐成为研究大时空尺度海洋现象的有效手段。自从Clarke等[53]开创利用遥感技术测定海面浮游植物叶绿素浓度以来,海洋初级生产力遥感已成为生物海洋学研究的一个重要课题。依据浮游生物的遥感数据,Platt等认为可用表层叶绿素估算水体叶绿素和初级生产力[54]。随着海洋调查规模的不断扩大和测定技术的多元化,简化海洋初级生产力数据的获取方法成为必须和可能。近年来荧光技术和遥测、遥感技术的发展使叶绿素的测定远比初级生产力的直接测定方便得多,通过测定叶绿素的含量来估算海洋初级生产力既简便又快速,特别适用于大范围的海洋调查。把生态学数理模型中的某些参数以遥感手段来获取,进行相应处理后用来估算海洋初级生产力,这是海洋初级生产力模型的主要形式,也是目前研究的热点。这类模型结合了浮游植物光合作用的生理学过程与经验关系,比较有代表性的模型有BPM(Bedford Productivity Model)模型[55, 56] 、LPCM(Laboratoire de Physique et Chimie Marines)模型[57]、VGPM(Vertically Generalized Production Model)模型[58, 59]等。
由上述研究可知,利用海水中叶绿素的含量估算海洋初级生产力时,首先要确定它们之间的换算系数,即同化系数。同化系数反映了植物光合色素的光合作用效率,是浮游植物光合作用能力的指标。在不同地区、不同季节、不同条件下,浮游植物的光合作用能力(同化系数)的变化很大,一般冬季的同化系数较低,夏末秋初较高。因此,无论是现场航测还是遥感数据分析,利用同化系数的年平均值或各海区的平均值来估算初级生产力,势必高估冬季的初级生产力,低估夏季和秋季的初级生产力。所以,在利用叶绿素估算海域初级生产力时,即使利用遥感技术测算海洋叶绿素含量,也需要现场同步测定调查海区的浮游植物的光合作用能力——同化系数。
3 真光层颗粒有机碳(POC)输出通量的估算方法研究真光层是海洋浮游生物活动最为活跃的区域,是海-气CO2交换的界面,是生源物质产生、再循环以及迁出到深海的一个重要水层,其向下输出的颗粒有机碳(POC)通量是衡量生物泵的运转效率的关键指标,并且决定着海洋颗粒活性元素和化学组分的生物地球化学循环速率。
海洋颗粒物质的传统采样方式是采集大量的海水过滤,要得到足够用于分析的颗粒物质样品,往往需要耗费大的工作量[60]。同时,由于海洋中颗粒物质的时空分布是不均匀的,因而无法取到反映海洋实况的样品。若要采集高分辨率的连续样品,需要科考船来回不断地奔波。另外,采用这种方法获得的样品包括了非自然沉降的颗粒,因而无法估计物质的净通量。20多年前,沉积物捕获器的发明,对于海洋现代生物地球化学过程研究具有划时代的意义。海洋沉积物捕获器大致有自由漂浮式、锚定式和中性浮力漂浮式沉积物捕获器等三类[61, 62, 63, 64]。
目前,海洋POC输出通量的研究主要基于两种方法:沉积物捕集器法和放射性同位素方法。利用沉积物捕获器通过单位时间、单位面积上收集到的颗粒物来定量POC输出通量。沉积物捕集器已被广泛用于测定深海的POC输出通量,方法可靠,结果准确。但在真光层中,由于水动力学、浮游动物等诸多因素的影响,以及沉降颗粒在捕集器内的溶解,由此方法获得的真光层POC输出通量一直备受海洋学家的质疑[61]。而中性浮动沉积物捕集器的问世和发展改善了水动力对捕集器的干扰问题,使得沉积物捕集器可以用来测定上层海洋的POC输出通量[62, 63, 64]。但这种捕集器设备造价昂贵,很难密集地布放于待研究海域,从而限制了这类沉积物捕集器的广泛应用。
天然放射性同位素示踪方法是测定POC输出通量的另一重要手段,应用最为广泛的是234Th—238U不平衡方法。234Th是一种天然的放射性核素,其半衰期为24.1 d。海水中的234Th是由238U(半衰期为 4.5×109a)不断进行α衰变产生的,它具有很强的颗粒活性,容易吸附在生源颗粒物上并随之沉降到深海,从而使它与母体238Th之间的放射性活度长期平衡被打破。通过测量真光层中234Th相对于238U的放射性活度比值,可得到234Th的输出通量,结合真光层底层颗粒物上有机碳与234Th的比值,可以得到从真光层底部输出的POC通量。用234Th法测量POC通量的优点在于可以得到颗粒物输出通量在几天到几周时间尺度上的平均值,且没有沉积物捕集器得到通量的明显偏差[65]。近几年来,随着海水234Th分析技术的不断发展,234Th—238U不平衡法在南大洋普里兹湾区[66]、太平洋的阿蒙森海区[67, 68]、大西洋威德尔海区冰架[69, 70]、中国南海[71, 72, 73, 74] 、中国台湾海峡[75, 76]、太平洋西北海域[77, 78]、大西洋地中海西北部[79, 80] 、北冰洋中部和西部海区[81, 82]等大洋和边缘海域的POC输出通量和颗粒动力学的研究中得到了广泛的应用,已被证明是研究上层海洋 POC输出通量的可靠方法。
在应用234Th—238U不平衡法研究海洋真光层POC输出通量时,颗粒物上的POC/234Th是制约POC输出通量估算准确性的一个重要因素。POC/234Th随采样地点和时间、浮游生物群落结构、颗粒粒径等变化而变化,其可以出现几个数量级的差别,这给POC输出通量的估算带来很大的误差[69, 77, 80, 83]。
4 海洋有机碳沉积通量的估算方法研究CO2从大气进入海洋后,在生物泵作用下形成颗粒有机碳并从上层水体输出到深层水体,大部分通过细菌分解作用转化为无机碳而可能重新返回大气层,只有很小一部分被埋藏在深海沉积物中长期封存,并在一定时间尺度上形成海洋碳汇作用的最终净效应,因此海洋有机碳沉积通量在碳循环研究中具有重要意义[84]。
海洋有机碳沉积 (SOC)通量测定需要先确定柱状沉积物的年龄,再结合表层沉积物的TOC得到有机碳沉积通量[85, 86]。放射性测年法是依据放射性元素蜕变等方法来测定地层年龄的方法。利用大气沉降到水及沉积物中的放射性核素(如210Pb、137Cs、14C等)的衰变定律,通过测量其放射性活度随深度的变化来计算沉积物的沉积速率,其适用的测年范围与所使用的放射性核素的半衰期有关[87]。在海底地层沉积物中应用较广的是230Th和210Pb法(半衰期分别为75 200 a和22.3 a),其中深海沉积速率和锰结核的生长速率主要用230Th法测定,浅海或近海松散沉积物多用210Pb法测定[88]。210Pb是238U系列中226Ra衰变中间产物222Rn的α衰变子体,半衰期为22.3a,属短寿命放射性同位素,被广泛用于百年时间尺度上的沉积物计年及沉积速率的测定,是研究近代江、河、湖、近海等沉积过程的重要手段。自然界中210Pb主要来源于地壳中238U的衰变和大气中210Pb的沉降,此外人工核反应也可产生210Pb。其中通过沉降并积蓄在沉积物中的210Pb因不与其母体共存和平衡,称为过剩210Pb (210Pbex)。210Pb测年法基于以下几点假设:(1)沉积体系为封闭系统,具备稳态条件;(2)沉降的210Pb能有效地转移到沉积物中,且不发生沉积后迁移作用;(3)沉积物中的非过剩210Pb与其母体226Ra保持平衡状态。虽然210Pb的沉降通量具有纬度效应,但同一地点210Pb的放射性通量在近百年的时间范围内可认为基本恒定[88, 89],沉积物中210Pbex的比活度将随沉积物质量深度呈指数衰减,因此对沉积物样品的210Pbex比活度分析,便可计算其沉积年龄。应用210Pb法进行海洋沉积物测年的过程中,根据沉积物的压实深度、沉积物的孔隙率、干沉积物的密度等参数确定沉积物中的210Pbex比活度衰变规律,算出某一深度的沉积物的年龄,结合210Pbex比活度随沉积物质量深度呈指数衰减的趋势,得到沉积物的沉积速率。根据沉积物中的有机碳含量即可得到有机碳沉积通量。
5 问题与展望海洋碳循环中海-气界面CO2交换通量、浮游生物的初级生产力、真光层POC输出通量和有机碳沉积通量间既有联系又相互区别,但目前的研究多限于单一过程中碳通量的研究,对上述过程的相互作用关系尚不明确,今后需加强碳在大气-海水-沉积物3种介质间交换通量之间相互影响的研究,提出海洋中碳垂直传输过程的主要影响因素和关键控制因子,并建立多元化的动态海洋碳通量分析系统,评估海洋不同时间尺度的碳汇效应,并与全球碳汇总量作比较,为研究海洋碳循环在全球碳循环中的作用提供客观参考依据。
此外,在海洋生态系统固碳估算方面,国内外还没有统一的规范和标准。我国应该把握此战略时机,通过室内模拟实验、船载模拟现场培养实验、海洋现场围隔实验,获取大量的数据资料和生态过程参数,并在此基础上建立有关数学模型,模拟不同环境条件和生态情景下的海洋碳循环物理泵和生物泵的调控机制和变动规律,特别是对我国陆架边缘海碳循环受控于多变的环境条件和复杂的调控机制。如大量陆源输入营养盐和上升流带来的营养盐,导致陆架海高的初级生产力。由于陆架海水较浅,形成的POC可以较快地到达海底沉积物中去,但浅水中的POC又会再悬浮,严重影响其生物泵的转化。通过上述研究,建立适合我国海洋固碳估算的整套观测技术、分析和计算方法,并构建海洋碳汇计算的指标体系、指标标准体系、以及评价标准体系等,服务于我国的碳“减排”、“增汇”国家需求。这方面研发也将产生巨大的生态环境效益、社会经济效益以及国际政治效应。 References:
| [1] | Solomon S, Qin D H, Manning M, Chen Z L, Marquis M, Averyt K B, Tignor M, Miller H L. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press, 2007: 996-996. |
| [2] | Baliño B M, Fasham M J R, Bowles M C. Ocean biogeochemistry and global change. IGBP Science, 2001, 2: 1-36. |
| [3] | Sarmiento J L, Monfray P, Maier-Reimer E, Aumont O, Murnane R J, Orr J C. Sea-air CO2 fluxes and carbon transport: a comparison of three ocean general circulation models. Global Biogeochemical Cycles, 2000, 14(4): 1267-1281. |
| [4] | Gloor M, Gruber N, Sarmiento J, Sabine C L, Feely R A, Rödenbeck C. A first estimate of present and preindustrial air-sea CO2 flux patterns based on ocean interior carbon measurements and models. Geophysical Research Letters, 2003, 30(1): 10-1-10-4. |
| [5] | Quay P D, Sommerup R, Westby T, Stutsman J, McNichol A. Changes in the 13C/12C of dissolved inorganic carbon in the ocean as a tracer of anthropogenic CO2 uptake. Global Biogeochemical Cycles, 2003, 17(1): 4-1-4-20. |
| [6] | Mikaloff-Fletcher S E, Gruber N, Jacobson A R, Doney S C, Dutkiewicz S, Gerber M, Follows M, Joos F, Lindsay K, Menemenlis D, Mouchet A, Müller S A, Sarmiento J L. Inverse estimates of anthropogenic CO2 uptake, transport, and storage by the ocean. Global Biogeochemistry Cycles, 2006, 20(2): GB2002. |
| [7] | Jacobson A R, Mikaloff-Fletcher S E, Gruber N, Sarmiento J L, Gloor M. A joint atmosphere-ocean inversion for surface fluxes of carbon dioxide: 1. Methods and global-scale fluxes. Global Biogeochemical Cycles, 2007, 21(1): GB1020. |
| [8] | Committee on Global Change. Toward an Understanding of Global Change. Washington, DC: National Academy Press, 1988: 56-56. |
| [9] | Liu H, Tang Q S. Review on worldwide study of ocean biological carbon sink. Journal of Fishery Sciences of China, 2011, 18(3): 695-702. |
| [10] | Yu W. Estimation and Determination of Carbon Fluxes on Three Interfaces of Western Arctic Ocean in Summertime. Beijing: Tsinghua University, 2010: 8-9. |
| [11] | Yin J P, Wang Y S, Xu J R, Sun S. Advances of studies on marine carbon cycle. Acta Ecologica Sinica, 2006, 26(2): 566-575. |
| [12] | Liss P S, Merlivat L. Air-sea gas exchange rates: Introduction and synthesis//Role of Air-Sea Exchange in Geochemical Cycling. Netherlands: Springer, 1986: 113-129. |
| [13] | Berger W H, Smetacek V S, Wefer G. Productivity of the Oceans: Present and Past. Chichester: John Wiley & Sons, 1989: 471-471. |
| [14] | Chen L Q, Yang X L, Zhang Y H, Li W, Lin Q, Lin H M, Xu S Q, Zhan J Q. Observation technology of CO2 fluxes in the ocean and atmosphere. Ocean Technology, 2008, 27(4): 9-12. |
| [15] | da la Paz M, Huertas E M, Padín X A, Gónzalez-Dávila M, Santana-Casiano M, Forja J M, Orbi A, Pérez F F, Ríos A F. Reconstruction of the seasonal cycle of air-sea CO2 fluxes in the Strait of Gibraltar. Marine Chemistry, 2011, 126(1/4): 155-162. |
| [16] | Li N, Li X G, Song J M. Key biogeochemistry processes of marine carbon cycle. Marine Environmental Science, 2005, 24(2): 75-80. |
| [17] | Matthews B J H. The Rate of Air-Sea CO2 Exchange: Chemical Enhancement and Catalysis by Marine Microalgae Type. Norwich: University of East Anglia, 1999: 68-79. |
| [18] | Chen Z X, Zhao Q. δ13C methods and its progress in the study of global carbon cycle. Advances in Earth Sciences, 2011, 26(11): 1225-1233. |
| [19] | McKinley G A, Follows M J, Marshall J, Fan S M. Interannual variability of air-sea O2 fluxes and the determination of CO2 sinks using atmospheric O2/N2. Geophysical Research Letters, 2003, 30(3): 1101-1104. |
| [20] | Lévy M, Lengaigne M, Bopp L, Vincent E M, Madec G, Ethé C, Kumar D, Sarma V V S S. Contribution of tropical cyclones to the air-sea CO2 flux: A global view. Global Biogeochemical Cycles, 2012, 26(2), doi: 10.1029/2011GB004145. |
| [21] | Bozec Y, Cariou T, Macé E, Morin P, Thuillier D, Vernet M. Seasonal dynamics of air-sea CO2 fluxes in the inner and outer Loire estuary (NW Europe). Estuarine, Coastal and Shelf Science, 2012, 100: 58-71. |
| [22] | Jouandet M P, Blain S, Metzl N, Mongin M. Interannual variability of net community production and air-sea CO2 flux in a naturally iron fertilized region of the Southern Ocean (Kerguelen Plateau). Antarctic Science, 2011, 23(6): 589-596. |
| [23] | Sejr M K, Krause-Jensen J, Rysgaard S, Srensen L L, Christensen P B, Glud R N. Air-sea flux of CO2 in arctic coastal waters influenced by glacial melt water and sea ice. Tellus B, 2011, 63(5): 815-822. |
| [24] | Chen L Q, Xu S Q, Gao Z Y, Chen H Y, Zhang Y H, Zhan J Q, Li W. Estimation of monthly air-sea CO2 flux in the southern Atlantic and Indian Ocean using in-situ and remotely sensed data. Remote Sensing of Environment, 2011, 115(8): 1935-1941. |
| [25] | Shadwick E H, Thomas H, Comeau A, Craig S E, Hunt C W, Salisbury J E. Air-sea CO2 fluxes on the Scotian Shelf: seasonal to multi-annual variability. Biogeosciences, 2010, 7(11): 3851-3867. |
| [26] | Boutin J, Merlivat L, Hénocq C, Martin N, SalléeJ B. Air-sea CO2 flux variability in frontal regions of the Southern Ocean from carbon Interface Ocean Atmosphere drifters. Limnology and Oceanography, 2008, 53(5): 2062-2079. |
| [27] | D'Ortenzio F, Antoine D, Marullo S. Satellite-driven modeling of the upper ocean mixed layer and air-sea CO2 flux in the Mediterranean Sea. Deep Sea Research Part I: Oceanographic Research Papers, 2008, 55(4): 405-434. |
| [28] | Huang Y S, Song J B, Fan C H. On the average time scale used in the calculations of the air-sea fluxes by eddy covariance method. Marine Sciences, 2011, 35(11): 114-119. |
| [29] | Hu Y B, Liu C Y, Yang G P, Zhang H H. The horizontal distributions of CO2 system parameters and sea-air flux in the North Yellow Sea and East China Sea in Autumn. Periodical of Ocean University of China, 2010, 40(2): 73-78. |
| [30] | Wanninkhof R. Relationship between wind speed and gas exchange over the ocean. Journal of Geophysical Research, 1992, 97(C5): 7373-7382. |
| [31] | Nightingale P D, Liss P S, Schlosser P. Measurements of air-sea gas transfer during an open ocean algal bloom. Geophysical Research Letters, 2000, 27(14): 2117-2120. |
| [32] | Riebesell U, Zondervan I, Rost B, Tortell P D, Zeebe R E, Morel F M M. Reduced calcification of marine plankton in response to increased atmospheric CO2. Nature, 2000, 407(6802): 364-367. |
| [33] | Fasham M J S, Ballifio B M, Bowles M C. A new vision of ocean biogeochemistry after a decade of the Joint Global Ocean Flux Study (JGOFS). AMBIO (Special Report), 2001, 10: 4-31. |
| [34] | González J M, Fernández-Gómez B, Fendàndez-Guerra A, Gómez-Consarnau L, Sánchez O, Coll-Lladó M, del Campo J, Escudero L, Rodríguez-Martínez R, Alonso-Sáez L, Latasa M, Paulsen I, Nedashkovskaya O, Lekunberri I, Pinhassi J, Pedrós-Alió C. Genome analysis of the proteorhodopsin-containing marine bacterium Polaribacter sp. MED152 (Flavobacteria). Proceeding National Academy of Science of the United States of America, 2008, 105(25): 8724-8729. |
| [35] | Suttle C A. Marine viruses-major players in the global ecosystem. Nature Reviews Microbiology, 2007, 5(10): 801-812. |
| [36] | Evans C, Thomson P G, Davidson A T, Bowie A R, Van den Enden R, Witte H, Brussaard C P D. Potential climate change impacts on microbial distribution and carbon cycling in the Australian Southern Ocean. Deep Sea Research Part Ⅱ: Topical Studies in Oceanography, 2011, 58(21/22): 2150-2161. |
| [37] | Bishop J K B, Wood T J. Year-round observations of carbon biomass and flux variability in the Southern Ocean. Global Biogeochemical Cycles, 2009, 23(2): 3206-3216. |
| [38] | Rivkin R B, Legendre L. Biogenic carbon cycling in the upper ocean: Effects of microbial respiration. Science, 2001, 291(5512): 2398-2400. |
| [39] | Williams P J L B. The balance of plankton respiration and photosynthesis in the open ocean. Nature, 1998, 394(6688): 55-57. |
| [40] | Seiter K, Hensen C, Zabel M. Benthic carbon mineralization on a global scale. Global Biogeochemical Cycles, 2005, 19(1): GB1010. |
| [41] | Song J M, Li X G, Yuan H M, Zheng G X, Yang Y F. Carbon fixed by phytoplankton and cultured algae in China coastal seas. Acta Ecologica Sinica, 2008, 28(2): 551-558. |
| [42] | Li B H, Fu K C, Hisayuki-Arakawa. Correlation between chlorophyll-a and primary productivity in the Southern Yellow Sea Area. Journal of Oceanography of Huanghai & Bohai Seas, 1998, 16(2): 48-53. |
| [43] | Ryther J H, Yentsch C S. The estimation of phytoplankton production in the ocean from chlorophyll and light data. Limnology and Oceanography, 1957, 2(3): 281-286. |
| [44] | Cadée G C, Hegeman J. Primary production of phytoplankton in the Dutch Wadden sea. Netherlands Journal of Sea Research, 1974, 8(2/3): 240-259. |
| [45] | Yentsch C S, Lee R W. A study photosynthetic light reaction, and a new interpretation of sun and shade phytoplankton. Journal of Marine Research, 1965, 24: 319-337. |
| [46] | Platt T, Gallegns C L, Harrison W G. Photoinhibition of photosynthesis in natural assemblages of marine phytoplankton. Journal of Marine Research, 1982, 38: 687-701. |
| [47] | Harrison W G, Plam T. Photosynthesis-irradiance relationships in polar and temperate phytoplankton populations. Polar Biology, 1986, 5(3): 153-164. |
| [48] | Yoshikawa T, Furuya K. Phytoplankton photosynthetic parameters and primary production in Japan Sea and the East China Sea: Toward improving primary production models. Continental Shelf Research, 2008, 28(7): 962-976. |
| [49] | Grant J, Bacher C, Cranford P J, Guyondetd T, Carreaue M. A spatially explicit ecosystem model of seston depletion in dense mussel culture. Journal of Marine Systems, 2008, 73(1/2): 155-168. |
| [50] | Zheng G X, Song J M, D J C, Wang Y M. Distributions of chlorophyll-a and carbon fixed strength of phytoplankton in autumn of the southern Huanghai Sea waters. Acta Oceanologica Sinica, 2006, 28(3): 109-118. |
| [51] | Fu M Z, Wang Z L, Li Y, Li R X, Lü R H, Sun P, Xia B. Study on size-fraction and carbon fixation of phytoplankton primary productivity in Jiaozhou Bay. Advances in Marine Science, 2009, 27(3): 357-366. |
| [52] | Xia B, Ma S S, Chen J F, Zhao J, Chen B J, Wang F. Distribution of organic carbon and carbon fixed strength of phytoplankton in Enteromorpha prolifera outbreak area of the Western South Yellow Sea, 2008. Chinese Journal of Environmental Science, 2010, 31(6): 1442-1449. |
| [53] | Clarke G L, Ewing G C, Lorenzen C J. Spectra of backscattered light from the sea obtained from aircraft as a measure of chlorophyll concentration. Science, 1970, 167(3921): 1119-1121. |
| [54] | Platt T, Herman A W. Remote sensing of phytoplankton in the sea: surface-layer chlorophyll as an estimate of water-column chlorophyll and primary production. International Journal of Remote Sensing, 1983, 4(2): 343-351. |
| [55] | Longhurst A, Sathyendranatll S, Platt T, Carla C. An estimate of global primary production in the ocean from satellite radiometer data. Journal of Plankton Research, 1995, 17(6): 1245-1271. |
| [56] | Kampel M, Sathyendranath S, Platt T, Gaeta S A. Satellite estimates of phytoplankton primary production at santos bight, southwestern-south Atlantic: Comparison of algorithms//Proceedings of the 2009 IEEE International Geoscience and Remote Sensing Symposium. Cape Town: IEEE, 2009, 2: 286-289. |
| [57] | Antoine D, André J M, Morel A. Oceanic primary production: 2. Estimation at global scale from satellite (Coastal Zone Color Scanner) chlorophyll. Global Biogeochemical Cycles, 1996, 10(1): 57-69. |
| [58] | Behenfeld M J, Falkowski P G. Photosynthetic rates derived from satellite-based chlorophyll concentration. Limnology and Oceanography, 1997, 42(1): 1-20. |
| [59] | Tripathy S C, Ishizaka J, Siswanto E, Shibataa T, Minob Y. Modification of the vertically generalized production model for the turbid waters of Ariake Bay, southwestern Japan. Estuarine, Coastal and Shelf Science, 2012, 97: 66-77. |
| [60] | Chen B, Hu L M, Deng S G, Fan D J, Guo Z G. Organic carbon in surface sediments of the Bohai Bay, China and its contribution to sedimentation. Marine Geology & Quaternary Geology, 2011, 31(5): 37-42. |
| [61] | Buesseler K O. Do upper-ocean sediment traps provide an accurate record of particle flux? Nature, 1991, 353(6343): 420-423. |
| [62] | Valdes J R, Price J F. A neutrally buoyant, upper ocean sediment trap. Journal of Atmospheric and Oceanographic Technology, 2000, 17(1): 62-28. |
| [63] | Sampei M, Sasaki H, Forest A, Fortier L. A substantial export flux of particulate organic carbon linked to sinking dead copepods during winter 2007-2008 in the Amundsen Gulf (southeastern Beaufort Sea, Arctic Ocean). Limnology and Oceanography, 2012, 57(1): 90-96. |
| [64] | Miquel J C, Martín J, Gasser B, Rodriguez-y-Baena A, Toubal T, Fowler S W. Dynamics of particle flux and carbon export in the northwestern Mediterranean Sea: A two decade time-series study at the DYFAMED site. Progress in Oceanography, 2011, 91(4): 461-481. |
| [65] | Moran S B, Weinstein S E, Edmonds H N, Smith J N, Kelly R P, Pilson M E Q, Harrison W G. Does 234Th/238U disequilibrium provide an accurate record of the export flux of particulate organic carbon from the upper ocean? Limnology and Oceanography, 2003, 48(3): 1018-1029. |
| [66] | He J H, Ma H, Chen L Q, Xiang B Q, Zeng X Z, Yin M D, Zeng W Y. The investigation on particulate organic carbon fluxes with disequilibria between thorium-234 and uranium-238 in the Prydz Bay, the Southern Ocean. Acta Oceanologica Sinica, 2008, 27(2): 21-29. |
| [67] | Amiel D, Cochran J K. Terrestrial and marine POC fluxes derived from 234Th distributions and delta 13C measurements on the Mackenzie Shelf. Journal of Geophysical Research-Oceans, 2008, 113(C3): C03S06. |
| [68] | Zhou K, Nodder S D, Dai M, Hall J A. Insignificant enhancement of export flux in the highly productive subtropical front, east of New Zealand: a high resolution study of particle export fluxes based on 234Th: 238U disequilibria. Biogeosciences, 2012, 9(3): 973-992. |
| [69] | Baena A M R Y, Boudjenoun R, Fowler S W, Miquel J C, Masqué P, Sanchez-Cabeza J A, Warnau M. 234Th- based carbon export during an ice-edge bloom: Sea-ice algae as a likely bias in data interpretation. Earth and Planetary Science Letters, 2008, 269(3/4): 596-604. |
| [70] | van der Loeff M R, Cai P H, Stimac I, Bracher A, Middag R, Klunder M B, van Heuven S M A C. 234Th in surface waters: Distribution of particle export flux across the Antarctic Circumpolar Current and in the Weddell Sea during the GEOTRACES expedition ZERO and DRAKE. Deep Sea Research Part Ⅱ: Topical Studies in Oceanography, 2011, 58(25/26): 2749-2766. |
| [71] | Chen W F, Cai P H, Dai M H, Wei J F. 234Th/238U disequilibrium and particulate organic carbon export in the northern South China Sea. Journal of Oceanography, 2008, 64(3): 417-428. |
| [72] | Ma H, Zeng Z, He J H, Chen L Q, Yin M D, Zeng S, Zeng W Y. Vertical flux of particulate organic carbon in the central South China Sea estimated from 234Th-238U disequilibria. Chinese Journal of Oceanology and Limnology, 2008, 26(4): 480-485. |
| [73] | Wei C L, Lin S Y, Sheu D D, Chou W C, Yi M C, Santschi P H, Wen L S. Particle-reactive radionuclides (234Th, 210Pb, 210Po) as tracers for the estimation of export production in the South China Sea. Biogeosciences Discussions, 2011, 8(12): 3793-3808. |
| [74] | Ma H, Zeng Z, Yu W, He J H, Chen L P, Cheng J P, Yin M D, Zeng S. 234Th/238U disequilibrium and particulate organic carbon export in the northwestern South China Sea. Acta Oceanologica Sinica, 2011, 30(3): 55-62. |
| [75] | Wei C L, Tsai J R, Wen L S, Pai S C, Tai J H. Nearshore scavenging phenomenon elucidated by 234Th/238U disequilibrium in the coastal waters off Western Taiwan. Journal of Oceanography, 2009, 65(2): 137-150. |
| [76] | Wei C L, Tsai J R, Hou Y R, Wen L S, Sheu D D, Chou W C. Scavenging phenomenon elucidated from 234Th/238U disequilibrium in surface water of the Taiwan Strait. Terrestrial, Atmospheric and Oceanic Sciences, 2010, 21(4): 713-726. |
| [77] | Buesseler K O, Pike S, Maiti K, Lamborg C H, Siegelb D A, Trullc T W. Thorium-234 as a tracer of spatial, temporal and vertical variability in particle flux in the North Pacific. Deep Sea Research Part I: Oceanographic Research Papers, 2009, 56(7): 1143-1167. |
| [78] | Maiti K, Benitez-Nelson C R, Buesseler K O. Insights into particle formation and remineralization using the short-lived radionuclide, Thoruim-234. Geophysical Research Letters, 2010, 37(15): L15608. |
| [79] | Szlosek J, Cochran J K, Miquel J C, Masque P, Armstrong R A, Fowler S W, Gasser B, Hirschberg D J. Particulate organic carbon-234Th relationships in particles separated by settling velocity in the northwest Mediterranean Sea. Deep Sea Research Par Ⅱ: Topical Studies in Oceanography, 2009, 56(18): 1519-1532. |
| [80] | Evangeliou N, Florou H, Scoullos M. POC and particulate 234Th export fluxes estimated using 234Th/238U disequilibrium in an enclosed Eastern Mediterranean region (Saronikos Gulf and Elefsis Bay, Greece) in seasonal scale. Geochimica Et Cosmochimica Acta, 2011, 75(19): 5367-5388. |
| [81] | Cai P, Van Der Loeff M R, Stimac I, Nöthig E M, Lepore K, Moran S B. Low export flux of particulate organic carbon in the central Arctic Ocean as revealed by 234Th: 238U disequilibrium. Journal of Geophysical Research-Oceans, 2010, 115: C10037. |
| [82] | Yu W, Chen L Q, Cheng J P, He J H, Yin M D, Zeng Z. 234Th-derived particulate organic carbon export flux in the western Arctic Ocean. Chinese Journal of Oceanology and Limnology, 2010, 28(6): 1146-1151. |
| [83] | Cochran J K, Miquel J C, Armstrong R, Fowler S W, Masqué P, Gasser B, Hirschberg D, Szlosek J, Baena A M R Y, Verdeny E, Stewart G M. Time-series measurements of 234Th in water column and sediment trap samples from the northwestern Mediterranean Sea. Deep Sea Research Part Ⅱ: Topical Studies in Oceanography, 2009, 56(18): 1487-1501. |
| [84] | Golderg E D, Koid M. Rates of sediment accumulation in the Indian Ocean//Geiss J, Golderg E D, eds. Earth Sciences and Meteoritics. Amsterdam: North company, 1963: 90-102. |
| [85] | Zaborska A, Carroll J, Papucci C, Torricellic L, Carroll M L, Walkusz-Miotk J, Pempkowiak J. Recent sediment accumulation rates for the Western margin of the Barents Sea. Deep Sea Research Part Ⅱ: Topical Studies in Oceanography, 2008, 55(20/21): 2352-2360. |
| [86] | Carroll J, Zaborska A, Papucci C, Schironec A, Carrolla M L, Pempkowiak J. Accumulation of organic carbon in western Barents Sea sediments. Deep Sea Research Part Ⅱ: Topical Studies in Oceanography, 2008, 55(20/21): 2361-2371. |
| [87] | Wan G J. 210Pb dating for recent sedimentation. Quaternary Sciences, 1997, 3: 230-239. |
| [88] | Liu W, Chu F Y, Li Q, Han X B. Dating methods about bulk sediments in marine environment. Journal of East China Institute of Technology: Natural Science Edition, 2011, 34(3): 257-265. |
| [89] | Sun L, Jie D M, Pu L J. An overview on the application of 210Pb and 137Cs dating in the research of recent sediment accumulation rate of coastal zone. Progress in Geography, 2007, 26(2): 67-76. |
| [9] | 刘慧, 唐启升. 国际海洋生物碳汇研究进展. 中国水产科学, 2011, 18(3): 695-702. |
| [10] | 余雯. 夏季西北冰洋三界面碳通量的估算与测定 [D]. 北京: 清华大学, 2010: 8-9. |
| [11] | 殷建平, 王友绍, 徐继荣, 孙松. 海洋碳循环研究进展. 生态学报, 2006, 26(2): 566-575. |
| [14] | 陈立奇, 杨绪林, 张远辉, 李伟, 林奇, 林红梅, 许苏清, 詹建琼. 海洋-大气二氧化碳通量的观测技术. 海洋技术, 2008, 27(4): 9-12. |
| [16] | 李宁, 李学刚, 宋金明. 海洋碳循环研究的关键生物地球化学过程. 海洋环境科学, 2005, 24(2): 75-80. |
| [18] | 陈中笑, 赵琦. 全球碳循环研究中的δ13C方法及其进展. 地球科学进展, 2011, 26(11): 122 5-1233. |
| [28] | 黄艳松, 宋金宝, 范聪慧. 海气通量涡相关法计算中的时间尺度分析. 海洋科学, 2011, 35(11): 114-119. |
| [29] | 胡玉斌, 刘春颖, 杨桂朋, 张洪海. 秋季东海和南黄海表层海水CO2体系各参数分布及海- 气界面通量. 中国海洋大学学报: 自然科学版, 2010, 40(2): 73-78. |
| [41] | 宋金明, 李学刚, 袁华茂, 郑国侠, 杨宇峰. 中国近海生物固碳强度与潜力. 生态学报, 2008, 28(2): 551-558. |
| [42] | 李宝华, 傅克忖, 荒川久辛. 南黄海叶绿素a与初级生产力之间的相关分析. 黄渤海海洋, 1998, 16(2): 48-53. |
| [50] | 郑国侠, 宋金明, 戴纪翠, 王益鸣. 南黄海秋季叶绿素a的分布特征与浮游植物的固碳强度. 海洋学报, 2006, 28(3): 109-118. |
| [51] | 傅明珠, 王宗灵, 李艳, 李瑞香, 吕瑞华, 孙萍, 夏滨. 胶州湾浮游植物初级生产力粒级结 构及固碳能力研究. 海洋科学进展, 2009, 27(3): 357-366. |
| [52] | 夏斌, 马绍赛, 陈聚法, 赵俊, 陈碧鹃, 王芳. 2008年南黄海西部浒苔暴发区有机碳的分布特征及浮游植物的固碳强度. 环境科学, 2010, 31(6): 1442-1449. |
| [60] | 陈彬, 胡利民, 邓声贵, 范德江, 郭志刚. 渤海湾表层沉积物中有机碳的分布与物源贡献估算. 海洋地质与第四纪地质, 2011, 31(5): 37-42. |
| [87] | 万国江. 现代沉积的210Pb计年. 第四纪研究, 1997, 3: 230-239. |
| [88] | 刘伟, 初凤友, 李琦, 韩喜彬. 海洋环境中松散沉积物的测年方法. 东华理工大学学报: 自然科学版, 2011, 34(3): 257-265. |
| [89] | 孙丽, 介冬梅, 濮励杰. 210Pb、137Cs计年法在现代海岸带沉积速率研究中的应用述评. 地理科学进展, 2007, 26(2): 67-76. |